
Turcolana Argano & Pesce, 1980
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4170.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:E8EBF58A-A009-41A9-9BF2-585719646622 |
|
DOI |
https://doi.org/10.5281/zenodo.5665243 |
|
persistent identifier |
https://treatment.plazi.org/id/957587A3-463C-AD7B-FF55-37A59CB0F8FC |
|
treatment provided by |
Plazi |
|
scientific name |
Turcolana Argano & Pesce, 1980 |
| status |
|
Genus Turcolana Argano & Pesce, 1980 View in CoL
Turcolana Botosaneanu, Bruce & Notenboom, 1985: 412 View in CoL .– Botosaneanu et al., 1985: 205 –206.– Botosaneanu & Notenboom, 1989: 2 –4, 13–14; 1992: 221.
Typhlocirolana View in CoL p. p.: Strouhal, 1960: 245 –256.– Por, 1962: 247 –252.
Type species. Turcolana cariae Argano & Pesce, 1980 ; by monotypy. Type locality, well near Çetibeli , between Ula and Köycegiz, Mugla province, SW Anatolia, Turkey .
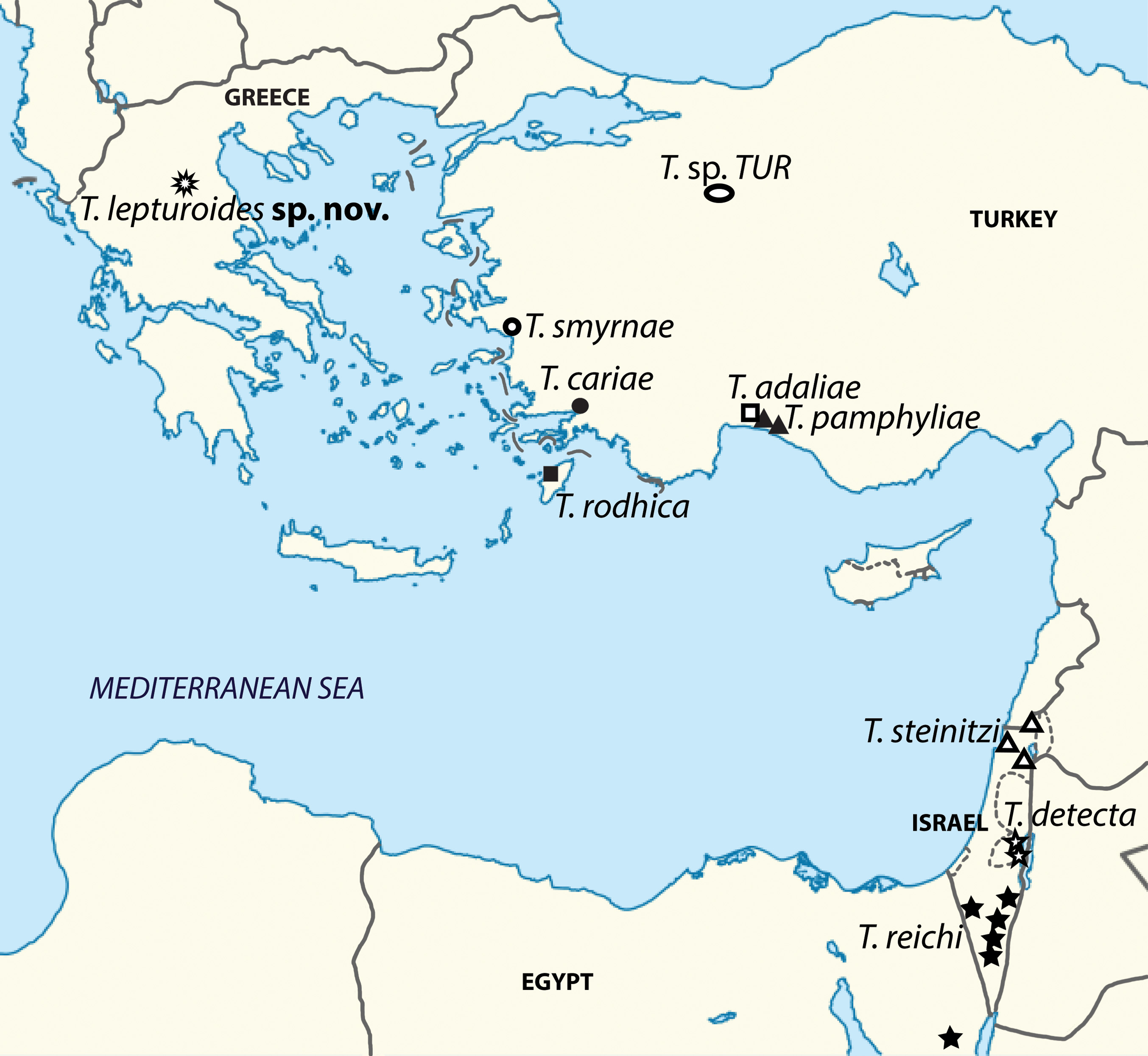
Species included. Turcolana rodhica Botosaneanu, Boutin & Henry, 1985 , from the underflow (hyporheal) of a spring brook in Ródos (Greek islands), T. steinitzi ( Strouhal, 1960) (= Typhlocirolana steinitzi Strouhal, 1960 ) from a water drill and springs in N Israel, T. reichi ( Por, 1962) (= Typhlocirolana reichi Por, 1962 ) from a small well in the Sinai Peninsula and wells and springs in S Israel, T. detecta Botosaneanu & Notenboom, 1992 , from small brackish springs on the west shore of the Dead Sea ( Israel), unveiled by small-scale regression of the sea, T. adaliae Botosaneanu and Notenboom, 1989 , from a well in Antalya province, SW Anatolia ( Turkey), T. pamphyliae Botosaneanu & Notenboom, 1989 , from the underflow of a river and a well in Antalya province, SW Anatolia ( Turkey), T. smyrnae Botosaneanu & Notenboom, 1989 , from the underflow of a river in Izmir province, SW Anatolia ( Turkey) and T. lepturoides sp. nov., described here.
Diagnosis (male). Depigmented and anophtalmic cirolanid. Body elongate, 3.3–4.2x as long as wide, convex. Appendage cuticular surfaces scale-like, consequently some margins serrate. Cephalon anterior margin smoothly rounded. Pereonite 1 and coxae 2–3 (pereonite epimera in previous references) each with posterolateral angle rounded; coxae 4–7 with distinct oblique carina, posterolateral margins posteriorly produced, posterolateral angle extending beyond (most species) or reaching ( T. reichi ) respective pereonites. Pleon with 5 visible unfused pleonites. Pleotelson apex never produced; posterolateral and posterior margins with short SS. Antennula (antenna 1) peduncle longer than flagellum; flagellum with 3–5 (most species), exceptionally 6 (single specimen of T. pamphyliae ) articles. Antenna (antenna 2) peduncle shorter than flagellum. All pereopods with secondary unguis on dactylus. Pereopod 1 subchelate, pereopods 2–7 ambulatory, increasing gradually in length posteriorly. Pleopod 1 largest and forming operculum; endopodite tapering, 1.3–1.4x as long and 1–1.2x as wide as exopodite; rami both with up to 7 PL. Pleopod 2 with appendix masculina inserted basally; endopodite 0.5–0.6x as long and 0.6–0.7x as wide as exopodite; rami both with up to 8 PL. Pleopods 3–5 endopodites shorter and narrower than exopodites, with 0 and up to7 PL, respectively; exopodites each with transverse suture. Uropods inserted ventrolaterally; sympodite posteromedial angle with 0–1 PL or medial margin with 1 PL and robust seta on posteromedial angle (only T. lepturoides , sp. nov.)
Description (male). Body about 3.3–4.2x as long as wide, with ability of at least strong curvation (most species) or complete volvation (alleged but not proven for T. cariae , T. rodhica ), Head 0.6–0.8x as wide as pereonite 1, enclosed laterally by pereonite 1. Body surfaces unornamented; pereonites 2–3 shortest, pereonites 5– 6 longest, pereonite 6 widest. Pleon with equally visible pleonites along dorsal margin, pleonite 5 (most species) and pleonite 4 ( T. pamphyliae , T. adaliae , T. rodhica ) narrowest and without epimera. Pleotelson with tendency towards medio-apical sinus, emargination or rounded; pleotelson lateral margins directed ventrally to form spacious pleonal cavity.
Antennula peduncle articles collinear, article 3 with large uniarticulate rounded lobe on distodorsal margin (not reported only in T. cariae ), interpreted either as scale (Wägele 1983, Brusca and Wilson 1991) or as fourth peduncular article ( Bruce 1986); article 1 short, article 2 longer, article 3 longest; flagellum with 3–5 (most species), exceptionally 6 articles (one specimen of T. pamphyliae ), about 0.7–0.75x (most species) or 0.55x ( T. lepturoides , sp. nov.) as long as peduncle, with 6 (most species) or 5 (some T. reichi ) aestethascs. Antenna peduncle articles 1–3 short, subequal, articles 4–5 long and slender; flagellum longer than peduncle, with 11–17 (most species) or 25–27 articles ( T. lepturoides , sp. nov.); antenna length about 0,3x (most species) up to 0,5x BL ( T. lepturoides , sp. nov.).
Frontal lamina protruding beyond or completely hidden behind median lobe of clypeus. Clypeus with differently protruding (strongly to moderately) median lobe of different shape (triangular, rounded).
Mandible incisor 3-cuspidate; molar process anterior margin with 10–27 teeth; spine row with 7–14 SR; palp with 3-articles, article 3 distally tapered. Maxilla with 3 endites; lateral and middle lobe subequal (most species) or middle lobe little shorter ( T. lepturoides , sp. nov.); lateral lobe with 2 PE; middle lobe with 3 PE; mesial lobe with numerous setae. Maxilliped endite with 4–6 setae and 1 coupling hook (most species) or 9 setae and 2 coupling hooks ( T. lepturoides , sp. nov.).
Pereopod 1 dactylus haptorial; merus inferior margin with single row of molariform CS and acute SRC; carpus inferior distal angle with single SRC and single SS. Pereopods 2–7 slender; basis superior margin with 2–5 slender plumose setae; merus inferior margin without molariform CS; pereopods 5–7 merus, carpus and propodus inferior margins with 0–3 SRC (most species) or more than 4 SRC ( T. lepturoides , sp. nov.); pereopods 2–3 with ( T. adaliae , T. steinitzi , T. reichi , T. detecta , T. lepturoides ) or without propodial organs.
Pleopod 1 sympodite rectangular; tapering endopodite with 3–7 PL and exopodite with 2–6 PL. Pleopod 2 appendix masculina moderately curved, slender, distally acute, 2–2.9x as long as endopodite; endopodite and exopodite with 1–8 and 2–6 PL, respectively. Pleopods 3–5 shape subequal, size increasing posteriorly; endopodites shorter than exopodites, with 0 and 2–7 PL, respectively. Uropods extending beyond pleotelson; endopodite medial margin with 0–1 SRC (most species) or 5–6 SRC ( T. lepturoides , sp. nov.).
Female. Non-ovigerous females are similar to males but for the sexual characters: pleopod 2 without appendix masculina. Pereopods 2–3 without propodial organs.
Remarks. Argano & Pesce (1980) emphasized the extraordinary modification of the uropods (i.e. large, flattened, completely withdrawing into the telson cavity), pleonite length ratio and the complex adaptation of the body of rolling into a perfect ball, as the most peculiar characters of the genus Turcolana , separating it from the west Mediterranean Typhlocirolana Racovitza, 1905 . While describing Turcolana rodhica , however, Botosaneanu et al. (1985) discarded lengths of pleonites and the distinctive shape of the uropods from the generic diagnosis due to the observed deviations and used “a terminal incisure receiving the cephalon when the body is rolled” as the sole character permitting the inclusion of the new species in the genus. Later, Botosaneanu & Notenboom (1989) disputed also the complete volvation as the alleged characteristic of all Turcolana species and prepared a list of characters distinguishing them from Typhlocirolana : a) stronger volvational tendency (vs. slight tendency towards curving the body), b) antennular flagellum with up to 6 articles (vs. with at least 7 flagellar articles), c) 1st pleopods larger than remaining ones, with tapering endopodite longer and wider or at most as wide as exopodite, rami both with reduced number of PL (vs. 1st pleopods not forming operculum, endopodite distally rounded, exopodite longer and wider than endopodite, both with numerous PL), d) uropod sympodite with at most one PL on medial margin (vs. mostly many but never less than 3 PL) and e) strongly ventrally directed lateral margins of pleotelson determining spacious pleonal cavity, pleotelson never forming medio-apical point and always with simple and short marginal setae (vs. less spacious pleonal cavity, pleotelson with medio-apical point, marginal setae long and plumose). Based on these characters, they reallocated Typhlocirolana steinitzi and T. reichi to the genus Turcolana . Just recently, another Typhlocirolana was reallocated, but the new genus Botolana Coineau & Boutin, 2015 was erected to accommodate Typhlocirolana leptura Botosaneanu, Boutin & Henry, 1985 .
Examination of the descriptions of Turcolana , Typhlocirolana and Botolana species, as well as the features of the new Turcolana , indicate that this genus can be defined by several unique characters, i.e. antennula flagellum mostly 4–5-articulate (exceptionally 6); pleopod 1 large and forming operculum, their endopodite tapering, much longer and wider or as wide as the exopodite, rami both with up to 7 PL; pleopod 3–5 endopodite without PL and exopodite with 2–7 PL; and uropod sympodite with less than 3 plumose setae on the medial margin. It has to be noted, however, that according to the drawings from the descriptions, all Turcolana species have 1 PL situated on the sympodite posteromedial angle (on the tip of the protuberance, not on the medial margin), except for the new species, with 1 PL situated a little above the tip, while on the tip, a robust seta is situated.
Other characters previously used in defining Turcolana , however, may not be shared by all members of the genus and/or may be shared by species from other two mentioned genera. For example, two ambiguous characters are: pleotelson apex not produced—also in Typhlocirolana tiznitensis Boulal, Boulanouar, Boutin & Yacoubi- Khebiza, 2009); and pleotelson only with short and simple marginal setae—also in Typhlocirolana tiznitensis and T. haouzensis Boutin, Boulanouar, Coineau & Messouli, 2002 .
Distribution and ecological notes. Turcolana is distributed around the eastern Mediterranean basin ( Fig. 1 View FIGURE 1 ), the Sinai Peninsula and central Greece representing the westernmost border of its distribution area. Based on the known distribution and the habitat preferences of Turcolana, Botosaneanu & Notenboom (1989) made several suppositions of its potential presence that may now be discarded. Due to the last finding in the cave stream of peninsular Greece, the potential distribution area of the genus may be considerably expanded as the potential localities seem to be restricted neither to phreatic and hyporheic groundwater nor the areas close to the actual shores of the eastern Mediterranean (valid also for all three Levantine species: T. reichi , T. steinitzi and T. detecta ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Turcolana Argano & Pesce, 1980
| Prevorčnik, Simona, Konec, Marjeta & Sket, Boris 2016 |
Turcolana
| Botosaneanu 1989: 2 |
| Botosaneanu 1985: 205 |
Typhlocirolana
| Por 1962: 247 |
| Strouhal 1960: 245 |