
Tapinoma nigerrima var. ibericum Santschi 1925
|
publication ID |
https://doi.org/10.11646/zootaxa.5435.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:121D0891-6348-49DB-B96D-7EE0CC6E62D3 |
|
persistent identifier |
https://treatment.plazi.org/id/945A3D69-FF99-FFB7-8394-AFB8FAE5FD64 |
|
treatment provided by |
Plazi |
|
scientific name |
Tapinoma nigerrima var. ibericum Santschi 1925 |
| status |
|
Tapinoma nigerrima var. ibericum Santschi 1925 [ type investigation]
The species has been described from Pozuelo de Calatrava, Spain based on a male. Investigated was the type male labeled “ Type ”, “ Tapinoma nigerrimum Nyl. ibericum Sants. type ” [Santschi’s handwriting], “POZUELO La Fuente” and “ANTWEB CASENT 0911578”, NHM Basel. The type was remounted and the genital prepared in an in-situ position. The allocation of the type specimen to the corresponding species cluster was shown by Seifert et al. (2016) in an analysis of male characters .
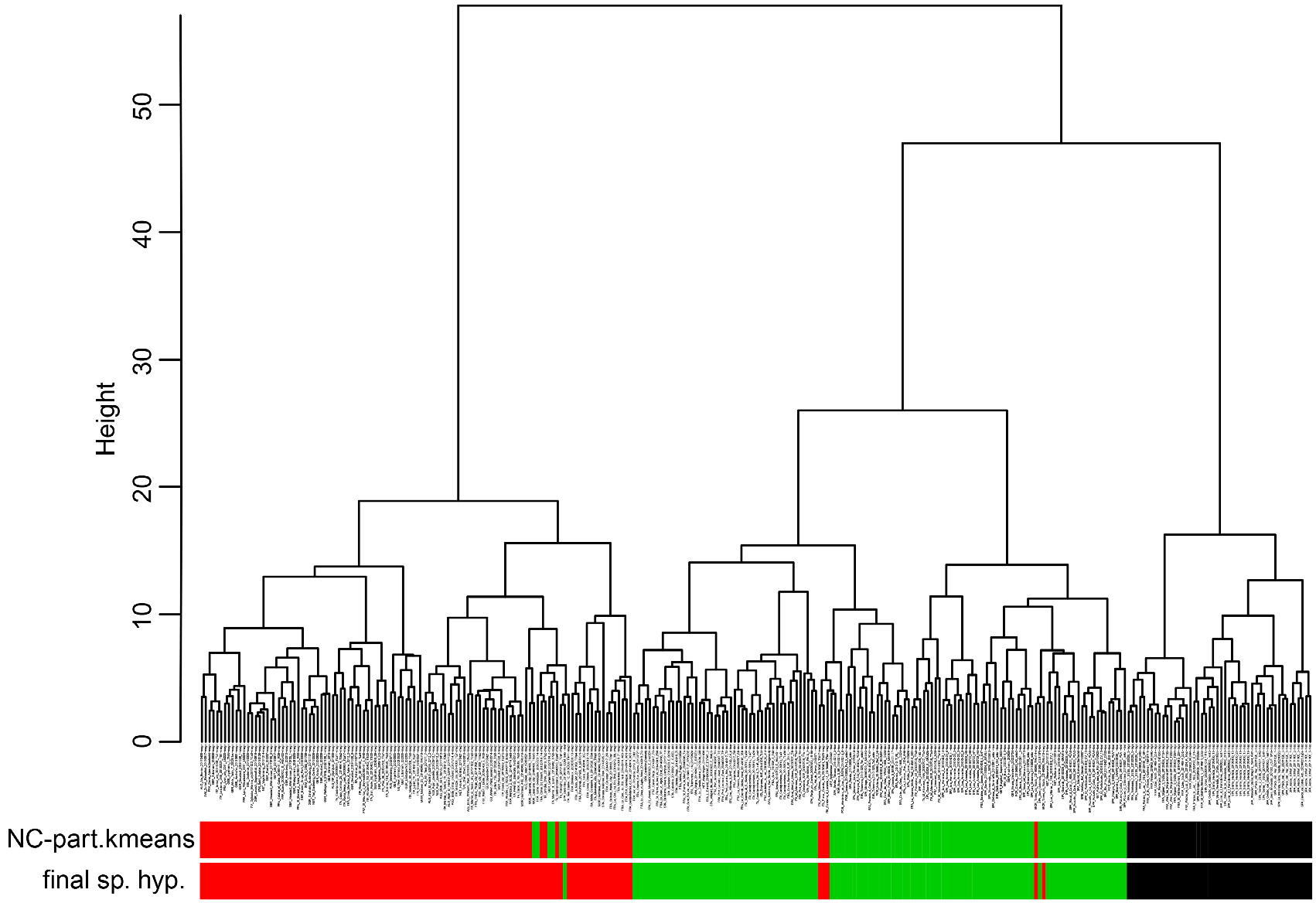
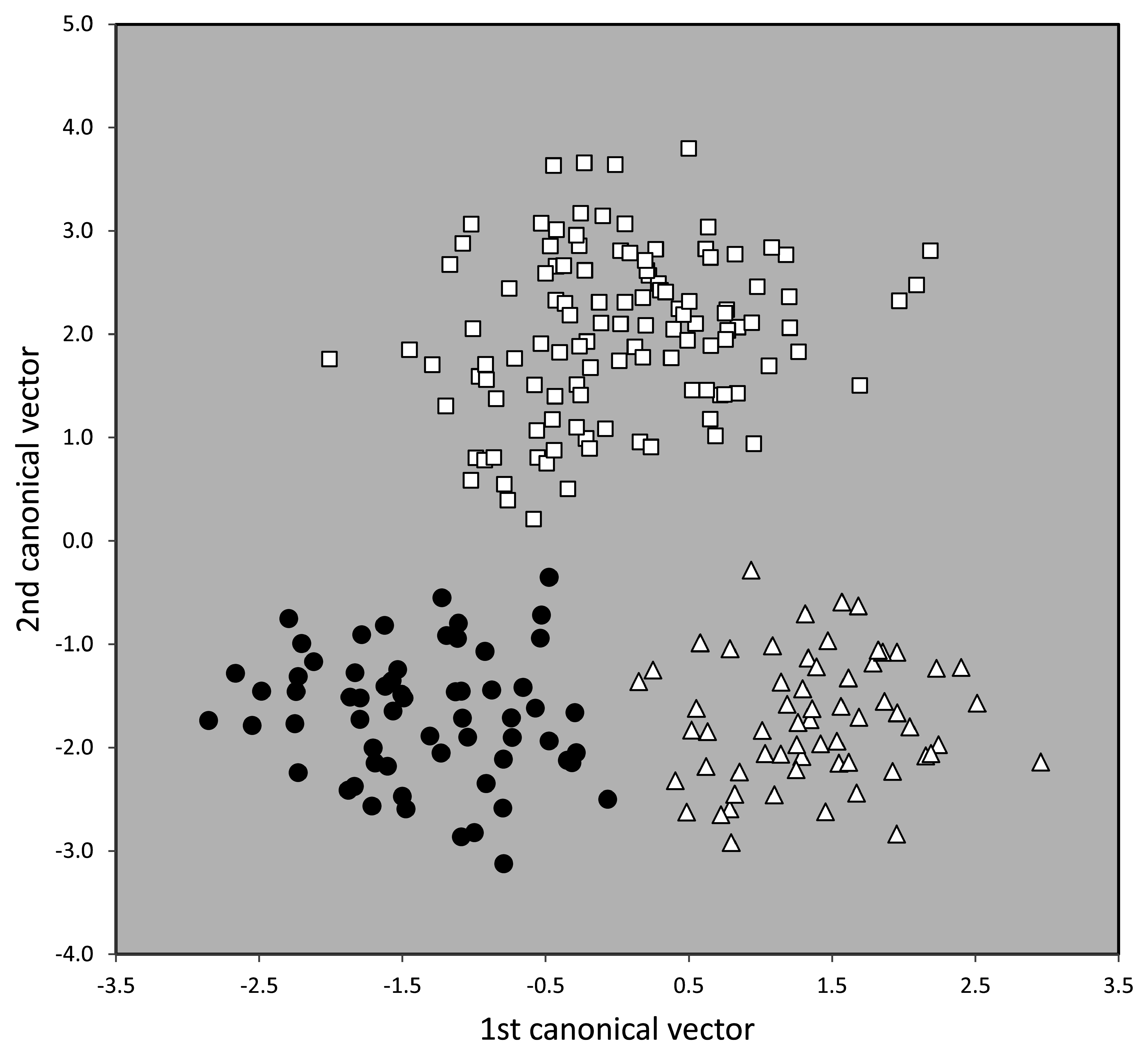
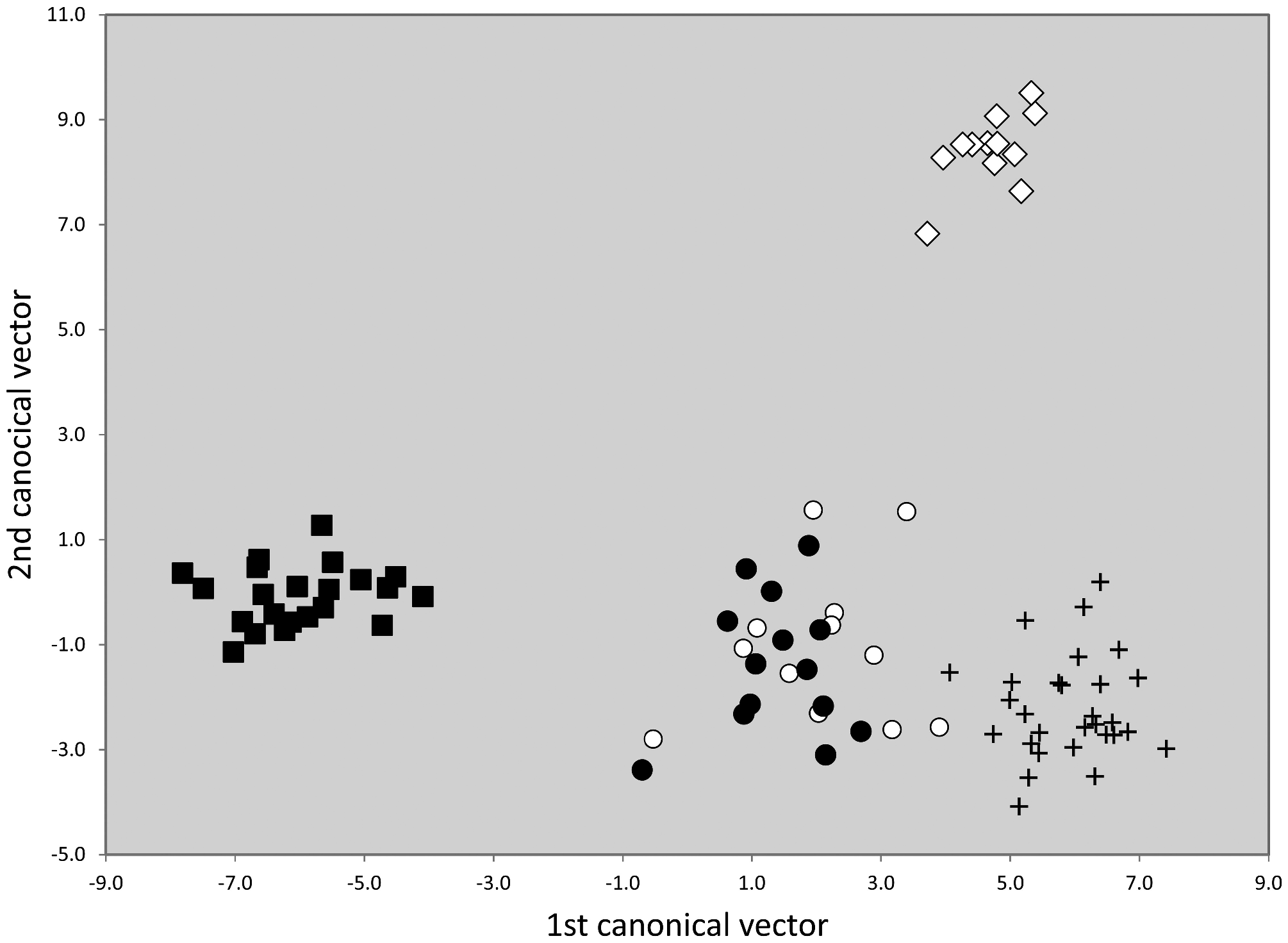
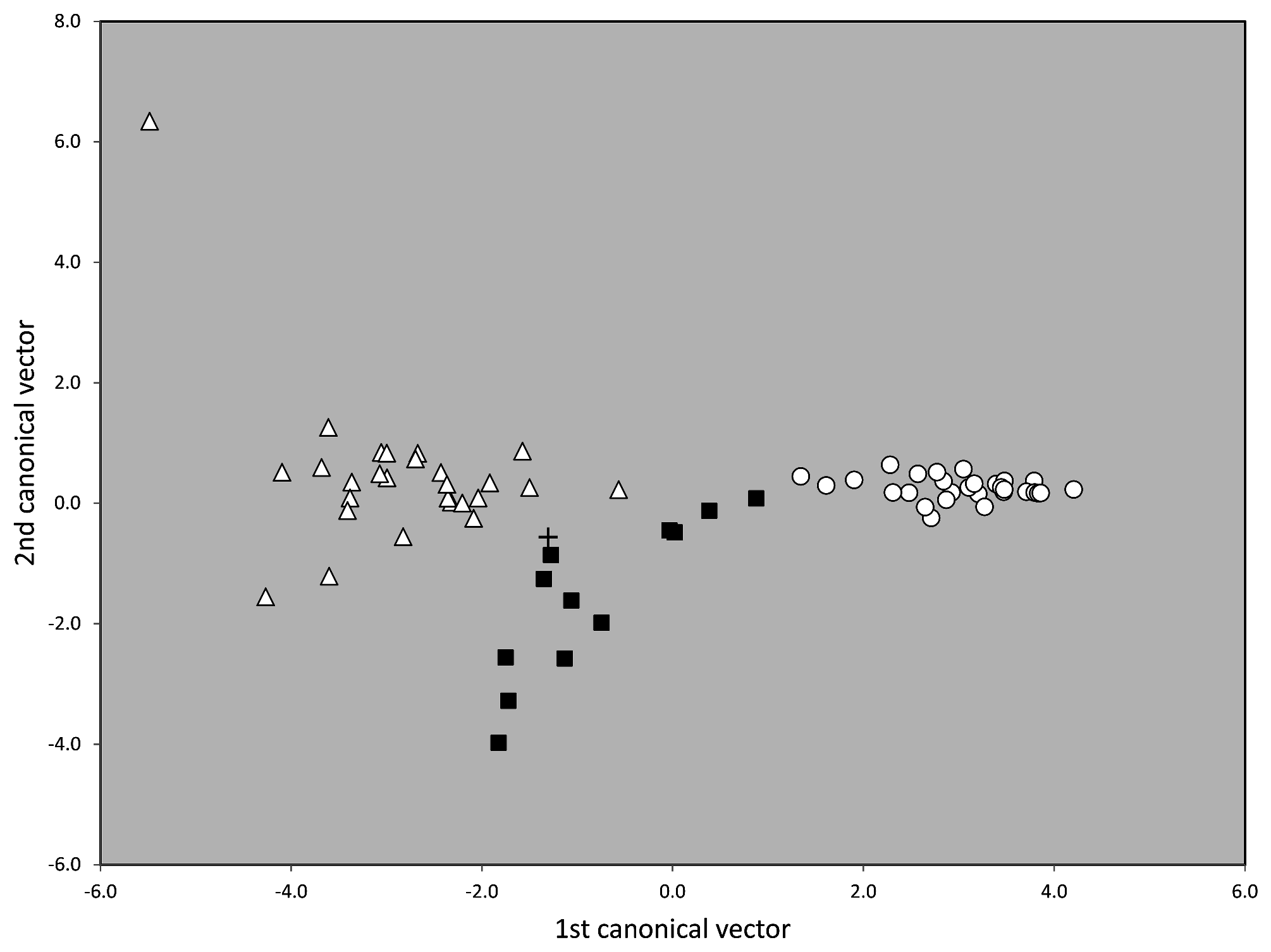
Material examined. Numeric phenotypical data were taken in 63 nest samples with 211 workers. They originated from France (1 sample), Germany (3), Great Britain (1), Netherlands (4), Portugal (6), and Spain (48). For details see supplementary information SI1, SI2 .
Geographic range. The natural range is apparently restricted to the Iberian Penisula. Populations north of the Pyrenees in France, Germany, the Netherlands and south England ( Isle of Wight) have been founded by anthropogenous introduction with plant material from Iberia. The northernmost known site, a supermarket in Burgdorf / Germany, is situated at 52.45°N and 10.00°E. Fifty-four nest sites in Iberia were situated at elevations of 623 ± 264 [7, 1264] m GoogleMaps .
Diagnosis:—Worker ( Tab. 3): All shape ratios given below are, in contrast to those in Tab. 3, primary ratios without RAV and all data are given as arithmetic mean ± standard deviation. Large, CS 977 ± 165 µm. Head broad CL/CW 1.032 ± 0.063. Postocular distance rather small and excavation of hind margin of vertex large, PoOc/CL 0.373 ± 0.010, ExOcc 2.25 ± 1.03%. Anteromedian clypeal excision deeper than in related supercolonial species and rather wide, ExCly/CS 10.47 ± 0.36%, ExClyW 6.53 ± 0.75%. The posterior, semicircular end of clypeal excision forms a concave plane delimited by a sharp ventral and a blunt dorsal edge. Sum of pubescence hairs and smaller setae protruding across the margin of clypeal excision including its dorsal edge very large, nExCly 18.80 ± 7.2. Scape moderately long, SL/CS 0.949 ± 0.047. Minimum distance of the inner margins of antennal socket rings smaller than in related supercolonial species, dAN/CS 0.292 ± 0.006. Eye moderately long, EL/CS 0.249 ± 0.016. Metanotal groove moderately deep, MGr/CS 3.25 ± 0.85 %. Mesosoma rather long and wide, ML/CS 1.273 ± 0.035, MW/CS 0.630 ± 0.019. Second funiculus segment shorter than in the supercolonial related species, Fu2L/CS 14.15 ± 0.53%, IFu2 1.898 ± 0.100. Pubescence, seta and pigmentation conditions as in T. magnum.
—Male ( Fig. 6 View FIGURE 6 ): the separation from the other species of the nigerrimum group by simple eye inspection of the genital appears difficult. However, the species can be separated by morphometry of the genital ( Seifert et al. 2014).
Taxonomic comments. The phenotypical and genotypical separation from other species of the T. nigerrimum group has been shown above ( Figs. 42 View FIGURE 42 , 44–46 View FIGURE 44 View FIGURE 45 View FIGURE 46 ).
Biology. Overall biology, effect on habitat, supercoloniality, interference potential and ecological impact in the introduction area is fully comparable to T. magnum. The lower total frequency of introduction to the north is probably less a question of specific biology but more a result of the geographic spacing of the trade lines leading to invasion of preferentially only NW Europe. In northern areas of introduction, it is not only an outdoor species but also occurring as nuisance in buildings. The nest construction is largely subterranean and often very extended. Nest entrances typically develop to big crater-like domes of ejected soil particles. Mounds of soil particles may be constructed at spots with a higher herb layer. Alates were observed in the nests 15 May ± 28 d [24 April, 15 June] n = 5. Knowledge on foraging is sparse. Portuguese populations began to forage in February at air temperatures of 10 °C. Morris et al. (2002) showed that T. ibericum was main predator of the Olive Moth Prays oleae (Bern.) in southern Spain but praying on egg-parasitoids of this moth may cause an opposite economic effect ( Pereira et al. 2004). Alvarez et al. (2023) suggested that it may potentially contribute to control olive pests without posing a risk to other natural enemies because it is usually not a hyper-predator. Yet, the role of T. ibericum differs between the crop species and depending on local conditions. Major damage by a huge supercolony (SaNo 579) is documented in a 200-ha walnut plantation in Herdade da Toula / Portugal. According to B. Leblanc (pers. comm. 2021) the damage was performed by (1) eating buds of young trees, (2) injuring young sprouts of older trees and imbibing their sap, (3) reducing pollination of fruits by eating pollen and by walking on flowers (which prevents the pollen to penetrate the pistil correctly), (4) fostering aphids, (5) killing predators of aphids and (6) extinguishing other ants.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |