
Tapinoma darioi Seifert et al. 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5435.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:121D0891-6348-49DB-B96D-7EE0CC6E62D3 |
|
persistent identifier |
https://treatment.plazi.org/id/945A3D69-FF98-FFA8-8394-AF9AFC4DFB50 |
|
treatment provided by |
Plazi |
|
scientific name |
Tapinoma darioi Seifert et al. 2016 |
| status |
|
Tapinoma darioi Seifert et al. 2016 [type investigation]
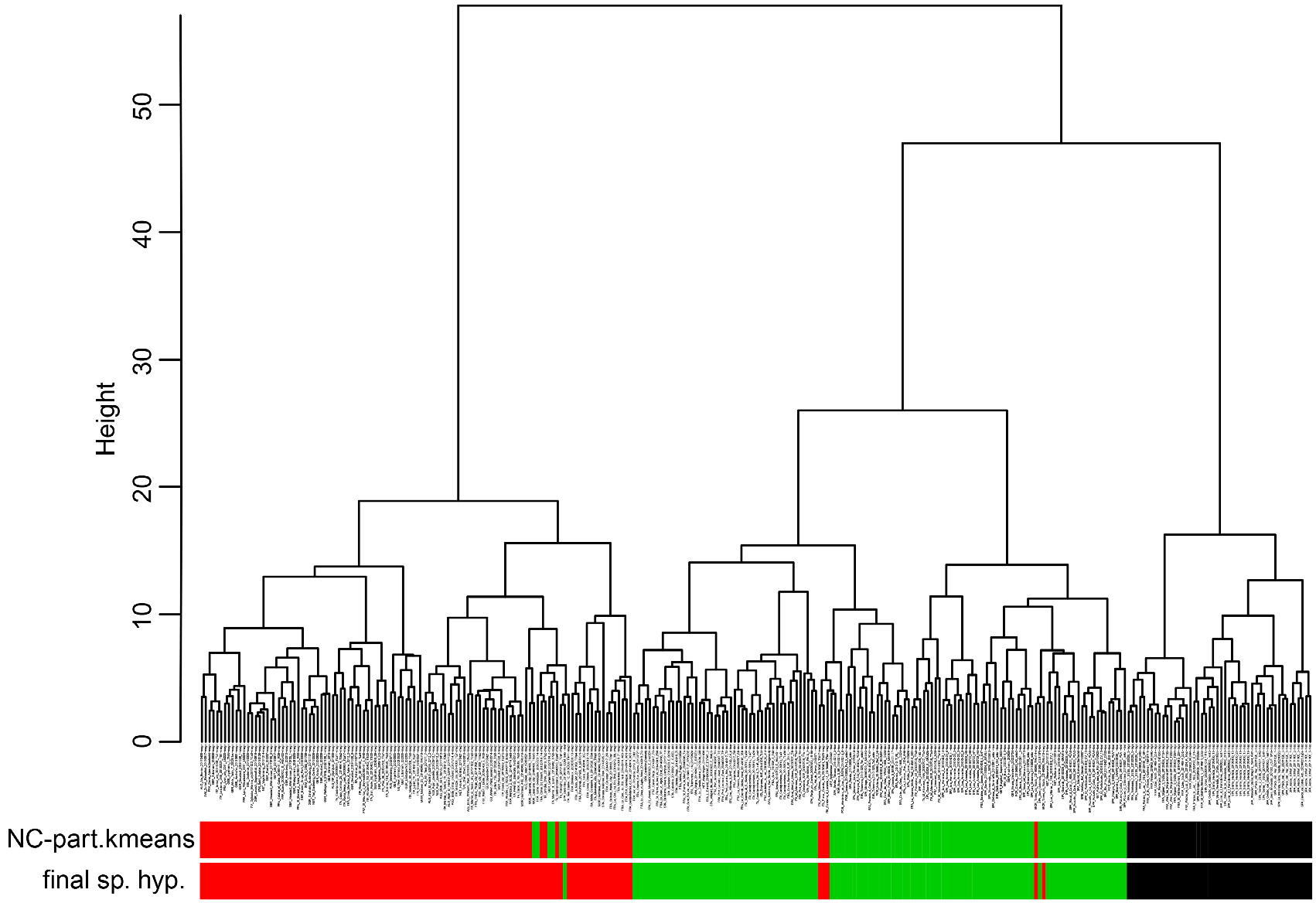
The species has been described from Rome, Italy. Investigated was the holotype, a major worker on the same pin with three paratype workers, labelled “ITA: 41.69858° N, 12.34985° E, Roma, Castelporziano, 1 m, Grotta di Piastra, dune, D’Eustacchio 20140423 - TLa45”, “ Holotype (top specimen) and paratypes Tapinoma darioi SEIFERT & al.”; two paratype gynes and two paratype males on other pins with the same collecting data; all material stored in SMN Görlitz. The allocation of the type specimens to the corresponding species cluster was shown by Seifert et al. (2016). In the extended present material, a wild-card run in a 5-class LDA considering the five morphologically separable entities of the T. nigerrimum species complex allocated the holotype sample with p=0.9873 to the T. darioi cluster GoogleMaps .
Material examined. Numeric phenotypical data were taken in 61 nest samples with 210 workers. They originated from France (24 samples), Italy (13), Netherlands (1) and Spain (23). For details see supplementary information SI1, SI2 .
Geographic range. According to the present state of information, Tapinoma darioi is continuously distributed along coastal areas (or near-coastal areas) of the Mediterranean Sea from Castellon (40.0°N, 0.0°E) northeast over Catalonia, the whole French coast east to Toulon, the Tyrrhenian coast of Italy from about 44.5°N south to 41°N, the Balearic Islands and Corsica. Three findings in Spain south of 37.5°N are far outside the main distribution range and certainly introductions. The only known introduction sites with established populations north of the Mediterranean zone are Limonest (45.82°N, 4.77°E), Chalon Sur Saone (46.78°N, 4.86°E), Illzach (47.77°N, 7.39°E) and Wageningen (51.98°N, 5.67°E). It is the T. nigerrimum group species occurring in the lowest altitudes: 61 findings range 80 ± 169 [0, 722] m, with 92% of all findings below 400 m. As a rule, occurrence more than 15 km away from the coast is rather rare in France ( Centanni et al. 2022) but is more often observed in Catalonia.
Diagnosis:—Worker ( Tab. 3): All shape ratios given below are, in contrast to those in Tab. 3, primary ratios without RAV and all data are given as arithmetic mean ± standard deviation. Smallest species of the T. nigerrimum group, CS 897 ± 135 µm. Head broad CL/CW 1.052 ± 0.052. Postocular distance rather small and excavation of hind margin of vertex large, PoOc/CL 0.380 ± 0.010, ExOcc 2.07 ± 0.85%. Anteromedian clypeal excision large and rather wide, ExCly/CS 9.71 ± 0.93%, ExClyW 6.14 ± 0.74%. The posterior, semicircular end of clypeal excision forms a concave plane delimited by a sharp ventral and a blunt dorsal edge. Sum of pubescence hairs and smaller setae protruding across the margin of clypeal excision including its dorsal edge large, nExCly 14.43 ± 5.7. Scape moderately long, SL/CS 0.965 ± 0.037. Minimum distance of the inner margins of antennal socket rings medium-sized, dAN/CS 0.300 ± 0.007. Eye moderately large, EL/CS 0.253 ± 0.014. Metanotal groove moderately deep, MGr/CS 2.80 ± 0.83 %. Mesosoma rather long and wide, ML/CS 1.274 ± 0.031, MW/CS 0.632 ± 0.021. Second funiculus segment longer than in the supercolonial related species, Fu2L/CS 14.50 ± 0.44%, IFu2 1.961 ± 0.106. Pubescence, seta and pigmentation conditions as in T. magnum.
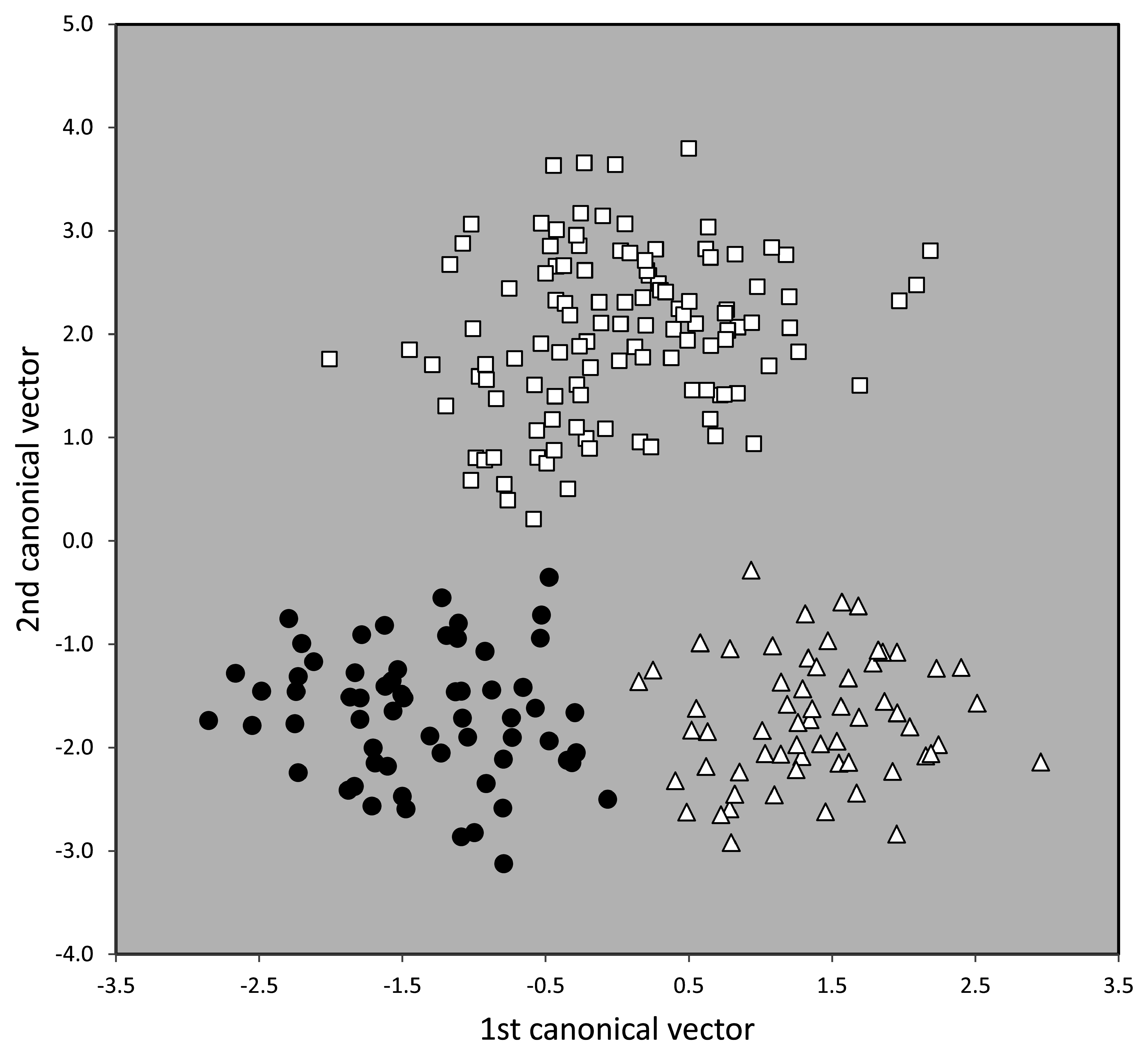
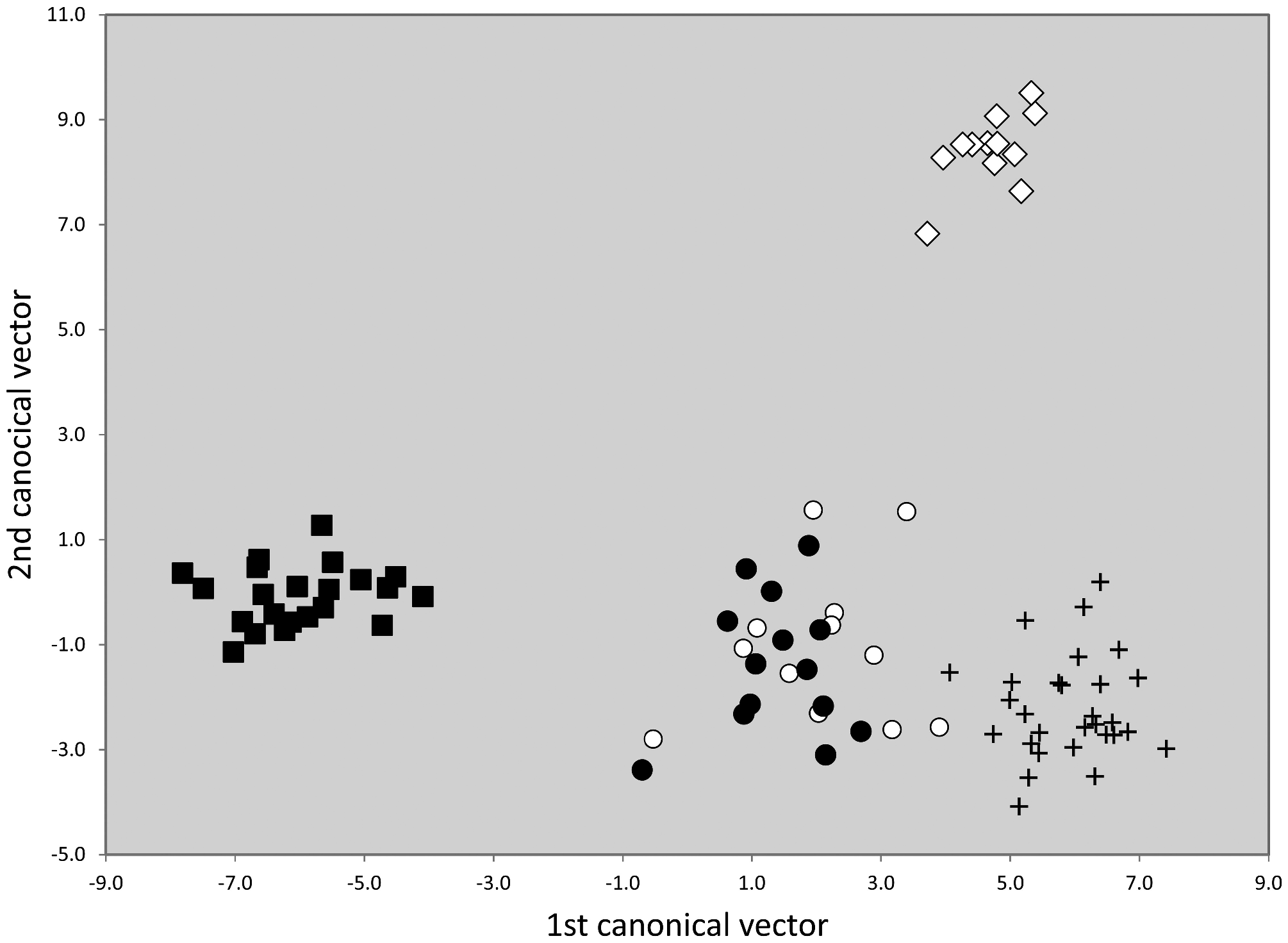
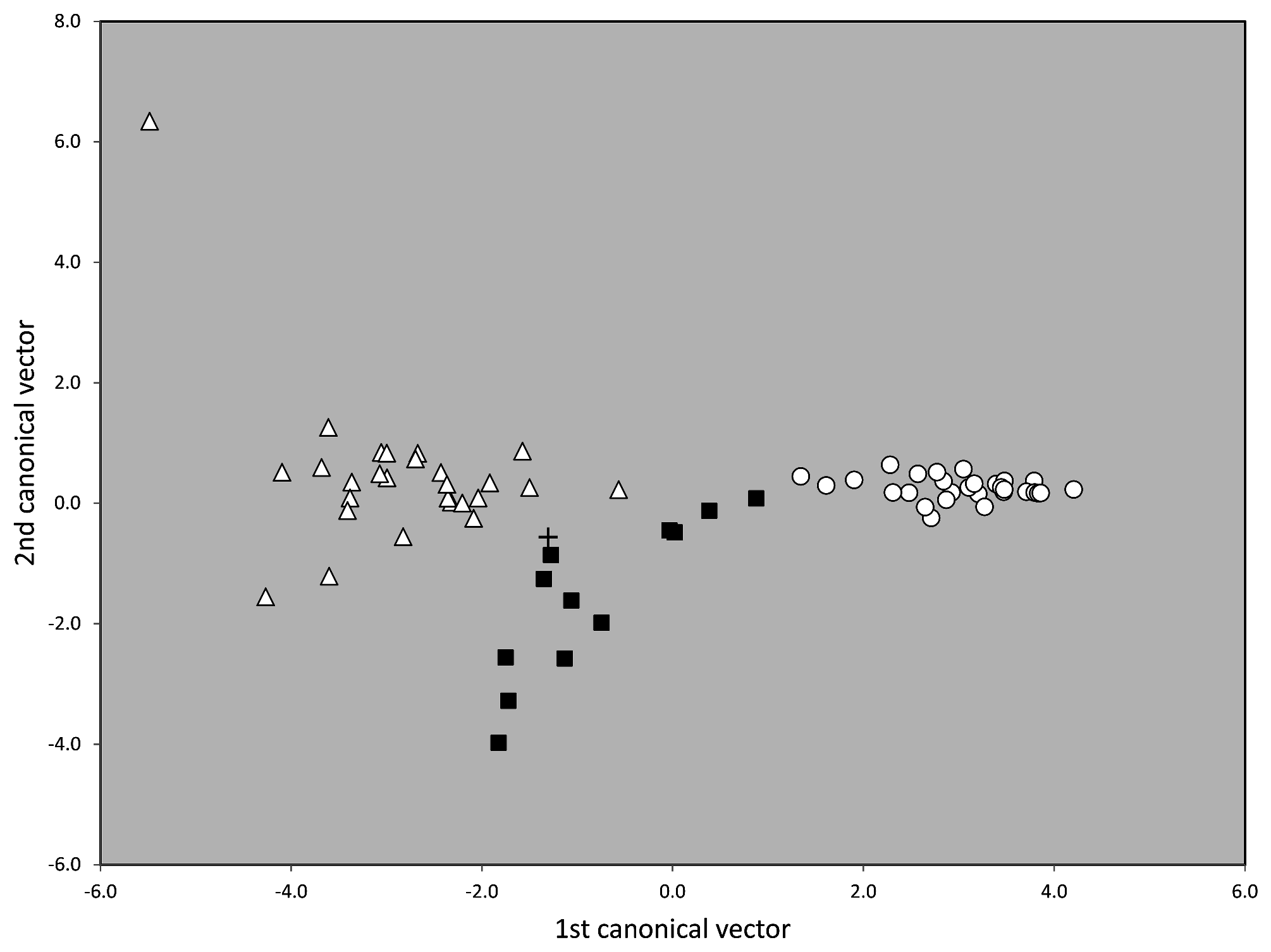
Taxonomic comments. The phenotypical and genotypical separation from other species of the T. nigerrimum group has been shown above ( Figs. 42 View FIGURE 42 , 44–46 View FIGURE 44 View FIGURE 45 View FIGURE 46 ).
Biology. Typical habitats are open unstable or degraded areas with very strong anthropogenic influence and a weakly developed tree layer but, in difference to T. magnum more frequently occurring in natural or semi-natural sand dunes. Niche separation is evident in Castelporziano Presidential Estate / Italy where T. darioi solely occurs on dunal systems while T. magnum is mainly linked to the Mediterranean scrub in much structured environments (D’Eustaccio et al. 2019). The nest construction is subterranean and often very extended, frequently reaching to a depth of 1 m. Nest entrances typically develop to big crater-like domes of ejected soil particles. Density data are missing. In the Mediterranean it is locally a eudominant species accounting for 95% of ants in some sites. Extremely polygynous and polydomous resulting in large supercolonies. The lower frequency of anthropogenous introduction in the Benelux countries and Germany is probably no question of specific biology but a result of the geographic spacing of the shipping of goods. It was observed in S France to limit the spread of the invasive Linepithema humile ( Blight et al. 2010) .
Alates were observed in the nests 22 May ± 20 d [23 Apr, 23 Jun] n = 12. The age and role of sexuals occasionally observed in September is unknown. Mating is probably intranidal or within the area of the supercolony. Foraging and nutrition was studied by ( Xerda et al. 1989) in lowland Spain. Foraging is diurnal from February to May and crepuscular and nocturnal from June to November. Very extensive trophobiosis with aphids occurs both on high trees, in the herb layer and the rhizophere; a permanent, 24-hour presence at aphid colonies was observed in less sun-exposed habitat spots. Honeydew accounts for probably>80% of total food mass. Nectarivory, though frequently observed with different plants, provides a very low contribution to energetic input. Plant parts are not consumed with the exception of elaiosomes of diverse myrmecochorous plants (e.g., Centaurea aspera , Euphorbia characias ) and of fallen sweet fruits. Carnivory of insects, snails and spiders is significant (perhaps 15% of total food mass) and probably more a collecting of carcasses than active hunting. Predation of sexuals of ants alighting after nuptial flight may be intensive. The mean and maximum size of solid food items retrieved by single workers are 2.6 and 6 mm. After discovery of very large food items mass recruitment starts, the item is disintegrated and the cut-off pieces are retrieved by single workers. Cooperative transport of items is so far unknown. Tool use: occasionally small twigs or pebbles are dropped into liquid food and retrieved after impregnation. In northern areas of introduction, it is not only an outdoor species but also occurring as nuisance in buildings.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |