
Coendou vestitus Thomas, 1899
|
publication ID |
https://doi.org/ 10.1093/mspecies/seab005 |
|
publication LSID |
lsid:zoobank.org:pub:EF3F6280-3468-455F-94B9-F3F847F85C79 |
|
persistent identifier |
https://treatment.plazi.org/id/941587DF-7608-9373-64BC-16305452EF03 |
|
treatment provided by |
Felipe |
|
scientific name |
Coendou vestitus Thomas, 1899 |
| status |
|
Coendou vestitus Thomas, 1899 View in CoL
Brown Hairy Dwarf Porcupine
Coendou vestitus Thomas, 1899:284 View in CoL . Type locality “ Colombia.”
[ Coendu View in CoL ] vestitus Trouessart, 1904:514 View in CoL . Incorrect subsequent spelling of Coendou vestitus Thomas, 1899 View in CoL .
Coendou (Sphiggurus) vestitus: Tate, 1935:307 View in CoL . Name combination.
Coendou (Sphiggurus) vestitus vestitus: Cabrera, 1961:603 View in CoL . Name combination.
Sphiggurus vestitus: Honacki, Kinman, and Koeppl, 1982:572 View in CoL View Cited Treatment . Name combination.
Coendu vestitus Rodríguez-Mahecha, Alberico, Trujillo, and Jorgenson, 2006:37 . Incorrect subsequent spelling of Coendou vestitus Thomas, 1899 View in CoL .
CONTEXT AND CONTENT. Context as for genus. No subspecies are recognized ( Voss 2015).
NOMENCLATURAL NOTES. Coendou vestitus Thomas, 1899 is the oldest name of a Neotropical porcupine with bristle– quills (Voss and da Silva 2001). The species was described based on a single specimen purchased by the British Museum in 1854 (Holotype, BMNH [British Museum of Natural History] 54.6.26.1— Thomas 1899; Voss and da Silva 2001). Based on the presence of bristle–quills C. vestitus was assigned to the “ vestitus ” group together with C. pruinosus (Voss and da Silva 2001) . C. pruinosus has been considered a subspecies or a junior synonym of C. vestitus ( Cabrera 1961; Honacki et al. 1982); however, both species show morphological and cytochrome- b differences and they are currently considered separate species (Voss and da Silva 2001; Voss et al. 2013; Voss 2015). Common names of C. vestitus include brown hairy dwarf porcupine and blackish hairy dwarf porcupine (English); puercoespín enano marrón peludo, erizo, puercoerizo, puercoespín marron, and puercoespín pardo (Spanish); Porco-espinho anão peludo marrom (Portuguese); Schwarzbrauner Greifstachler (German); and Coendou brun (French— Rodríguez-Mahecha et al. 1995; Emmons and Feer 1997; Voss 2015; Barthelmess 2016).
DIAGNOSIS
Coendou differs from the other two representatives of the family Erethizontidae , the broomstraw-spined porcupine ( Chaetomys subspinosus ) and the North American porcupine ( Erethizon dorsatum ) by a combination of characters ( Voss 2015; Barthelmess 2016). Coendou can be differentiated from the North American porcupine by the presence of a robust tail exhibiting a well-developed hairless, prehensile surface (the North American porcupine has a short, thick tail that is completely covered in hair). Within Coendou , tail length varies depending on the species (from about 40% to> 100% of the head–body length— Voss 2011, 2015; Barthelmess 2016). The pelage of Coendou may include both quills and hairs, which can partially or completely cover the quills. The quills of Coendou are long, straight or slightly curving and in some parts of the body, like the rump, the spines are defensive with recurved barbs (quills with a unique shape, wavy or kinked, and barbless in the broomstraw-spined porcupine— Voss 2015; Barthelmess 2016; Chapman et al. 2019). The pinna in Coendou and the broomstrawspined porcupine is reduced to an angular ridge of almost naked skin (small but distinct and densely furred in the North American porcupine— Voss 2015). The mystacial vibrissae of Coendou and the broomstraw-spined porcupine are long and extend well behind the ear (short and scarcely reaching the base of the ear in the North American porcupine— Voss 2011). The skull of Coendou and the North American porcupine lack well-developed postorbital processes (well-developed in the broomstraw-spined porcupine). Cranial sutures in Coendou disappear with age (except for sutures surrounding the premaxillae and the auditory bullae) and, in three species, the frontal sinuses are conspicuously inflated. The upper cheek teeth are low-crowned and wider than long ( Voss 2015).
Coendou vestitus can be distinguished from other species of Coendou ( C. bicolor , C. prehensilis , C. rufescens , and C. speratus ) by the dorsal pelage which includes defensive quills, bristle–quills, and long fur. Only three additional species of Coendou ( ichillus , pruinosus , and roosmalenorum ) have bristle–quills; however, these are bicolored in C. vestitus , and tricolored in the other species ( Voss 2011).
GENERAL CHARACTERS
Coendou vestitus ( Fig. 1 View Fig ) is a small-sized porcupine covered by three types of hair: soft fur (fine wavy hairs), bristle–quills (diameter <1 mm, length to 70 mm, mostly black with pale yellowish in the proximal 25 mm), and defensive quills (length 25–37 mm and diameter 1.2 mm), that are bicolored with pale yellow or white ivory in the basal portion and brown or blackish in their distal portion (Emmons and Feer 1997). Dorsal fur is abundant and relatively long (70–80 mm), yellowish to uniform brown, blackish, or warm smoky brown. This fur blends with bristle–quills, completely or partially covering the quills except on head and face, where white-tipped quills emerge through hair (Emmons and Feer 1997; Alberico and Moreno 2006). The underparts are covered with abundant soft brown hair, slightly lighter than the dorsum (Emmons and Feer 1997; Alberico et al. 2000; Voss and da Silva 2001; Alberico and Moreno 2006). The fur on the sides is shorter and more sparse but sufficient to hide the quills (Emmons and Feer 1997; Alberico et al. 2000). The snout carries long, black mystacial vibrissae. Also present are supraorbital (superciliary), genal (on the cheeks), and mandibular vibrissae. Additionally, C. vestitus exhibits postcranial vibrissae between the elbow and the wrist, between the knee and the ankle, and on the ventral surface of the body between the anterior and posterior limbs (Voss and da Silva 2001).
The dorsal part of the hindlegs is dark brown as is the dorsum. The tail presents bristle and defense quills in the proximal one-half; the tail tip is prehensile, naked, and callused (Emmons and Feer 1997; Alberico et al. 2000). The upper surfaces of the forefoot and hindfoot are densely covered with thick brown hairs like the color of the dorsum (Voss and da Silva 2001; Alberico and Moreno 2006).
Mean external measurements (mm; with parenthetical range and n) for five females and three specimens of unknown sex (not all measurements were available for all specimens) were: head–body length, 346 (330–370; 6); tail length, 179 (170–195; 6); length of hind foot, 60 (56–63; 6—Voss and da Silva 2001). Mean craniodental measurements (mm; with parenthetical range and n) for the same eight specimens mentioned previously (Voss and da Silva 2001) were: condylo-incisive length, 66.8 (65.4– 68.3; 6); length of diastema, 17.4 (16.9–17.9; 7); length of incisive foramen, 4.2 (3.6–4.9; 7); breadth of incisive foramina, 2.9 (2.7–3.1; 7); maxillary toothrow, 15.5 (14.7–16.4; 7); length of molars, 11.3 (10.8–12.0; 7); breadth of P4, 4.5 (4.1–4.9; 7); breadth of M1, 4.0 (3.8–4.3; 7); anterior palatal breadth, 4.9 (4.2–5.6; 7); posterior palatal breadth, 6.2 (5.1–7.2; 7); posterior zygomatic breadth, 42.9 (42.1–43.5; 6); height of the infraorbital foramen, 10.4 (9.8–11.0; 7); zygomatic length, 27.2 (26.2–28.1; 7); length of nasals, 20.2 (18.9–21.8; 5); breadth of nasal aperture, 10.7 (10.4–11.2; 7); breadth of braincase, 29.8 (27.8–30.6; 7); depth of upper incisor, 3.3 (3.1–3.5; 7); breadth of upper incisor tips, 4.6 (4.2–4.8;7—Voss and da Silva 2001; Voss 2015).
DISTRIBUTION
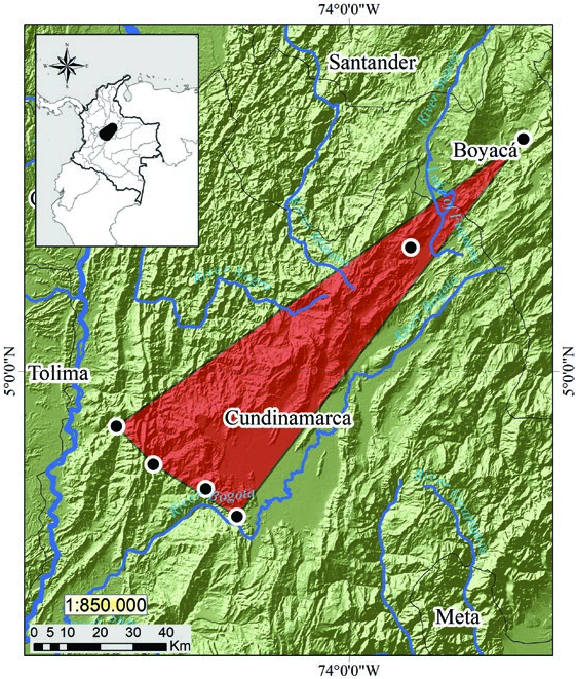
Coendou vestitus is an endemic Andean species restricted to the western foothills of the Eastern Cordillera of Colombia ( Fig. 2 View Fig ) in an elevational range from 1,300 to 2,890 m (Alberico and Moreno 2006; Voss 2011, 2015; Ramírez-Chaves et al. 2016). It has been recorded in six localities ( Ramírez-Chaves et al. 2019). Five confirmed localities are in the Department of Cundinamarca: 1) San Juan de Río Seco; 4°51 ′ 4 ″ N – 74°38 ′ 1 ″ W; 1,440 m (AMNH [American Museum of Natural History, New York] 70596, 71359; BMNH [Natural History Museum, London] 24.2.21.2; MNHN [Muséum National d’Histoire Naturelle, Paris] 1929.631, 1929.632; USNM [National Museum of Natural History, Washington, D.C.] 240035; MLS [Museo de La Salle] 248—Voss and da Silva 2001; Voss 2011); 2) Quipile; 4°44 ′ 50 ″ N – 74°31 ′ 59 ″ W; 1,250 m (AMNH 70529— Voss and da Silva 2001; Voss 2015); 3) Ubaté 5°20 ′ 14.72 ″ N – 73 °49 ′ 57.16 ″ W; 2,890 m (photograph— La Villa 2018); 4) Bogotá, Parque Natural Chicaque; 4°36 ′ 19.3 ″ N – 74°18 ′ 19.8 ″ W; 2,625 m (photograph— Ramírez-Chaves et al. 2019); 5) Tena, Laguna de Pedro Palo; 4°40 ′ 49 ″ N – 74°23 ′ 24.3 ″ W; 2,080 m (photograph— Ramírez-Chaves et al. 2019); and one in the Department of Boyacá, 6) Villa de Leyva; 5°37 ′ 59 ″ N – 73°31 ′ 32 ″ W; 2,070 m (IAvH [Instituto de Investigaciones Biológicas Alexander von Humboldt] 7956 and MLS 753— Weksler et al. 2016). Confirmed localities are located in the ecoregions Northern Andean Páramo and Magdalena Valley montane forests, that include the tropical dry forests and the montane ecosystems ( Ramírez-Chaves et al. 2019). One record from Department of Meta, Villavicencio: 4°10 ′ 0 ″ N – 73°39 ′ 0 ″ W; 610 m (ICN [Instituto de Ciencias Naturales, Universidad Nacional de Colombia] 3505— Alberico et al. 2000; Voss 2011; Ramírez-Chaves et al. 2016), located on the eastern side of the Eastern Cordillera is problematic, as the information contained on the specimen label indicates that the individual was purchased in a marketplace in Bogotá, but the origin was thought to be Villavicencio (Alberico and Moreno 2006). For that, the locality of the record “Villavicencio” is considered false ( Ramírez-Chaves et al. 2019). No fossils are known.
FORM AND FUNCTION
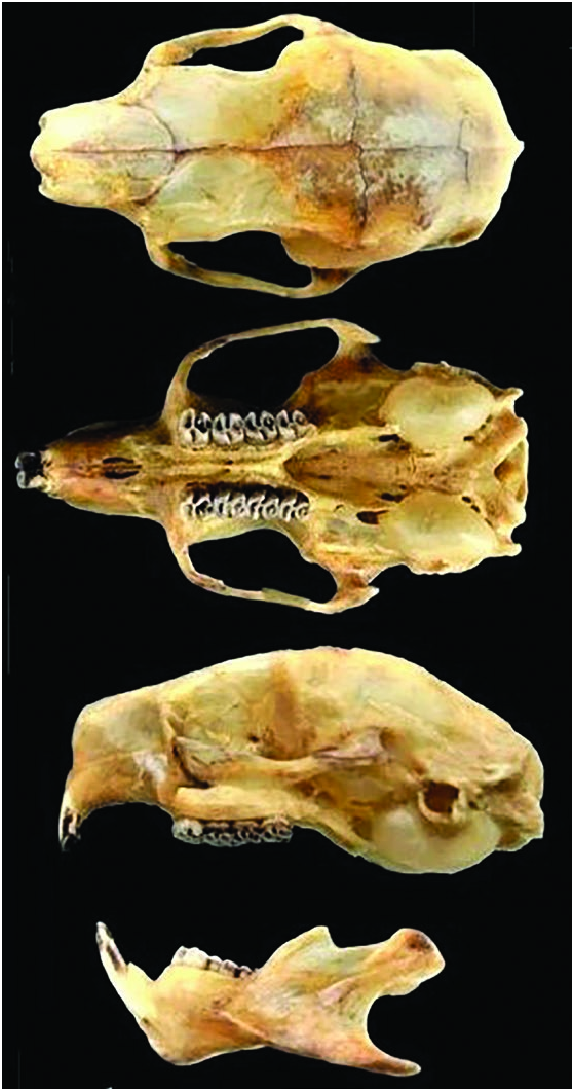
Form. —Frontal sinuses are not inflated in the adult skull
( Fig. 3 View Fig ) and the roof of the external auditory meatus is weakly developed (Voss and da Silva 2001). The rostrum is short and tapering in younger animals. The nasal bones are approximately parallel-sided, neither increasing nor decreasing posteriorly in breadth, with rounded posterior margins that extend well behind the premaxillae. Viewed from above, the zygomatic arches converge anteriorly from their widest point at the level of the squamosal roots with only a slight lateral deflection at the level of the orbits. The jugals are slender without a conspicuous postorbital expansion. The dorsolateral contours of the braincase have only a slight to moderate sculpting at the origin of the temporalis muscle; the left and right temporal scars are widely spaced and do not join middorsally to form a sagittal crest (Voss and da Silva 2001). The mesopterygoid fossa penetrates anteriorly to or between the posterior part of the second molars, and the bony roof of the fossa is not perforated by distinct sphenopalatine vacuities. The alisphenoid is completely ossified, resulting in a sphenopterygoid canal that opens laterally; the buccinator–masticatory foramen is confluent with the foramen ovale (Voss and da Silva 2001). The auditory bullae are small (ca. 14–15 mm), rounded capsules that are well-separated from the base of the paraoccipital process on each side. The dorsal roof of the external auditory meatus has an indistinct bony ridge that is less well-developed compared to that seen in some congeneric taxa (e.g., Coendou melanurus ). The mandible is distinctive in the absence of a well-defined coronoid process, which is represented only as a rounded convexity at the base of the ascending ramus in all specimens examined (Voss and da Silva 2001).
The small incisive foramina are completely contained in the premaxilla or shallowly contacting but not deeply penetrating between the maxilla; the left and right foramina are incompletely separated and are recessed in a common fossa in some specimens, but in others they are completely separated and are not recessed together. The posterior diastema is marked by shallow and widely separated lateral sulci. In all the specimens examined, the palatal bridge (between the toothrows) has a welldeveloped central keel and deep lateral gutters (Voss and da Silva 2001). The buccinator–masticatory foramen and foramen ovale are confluent (Voss and da Silva 2001; Voss 2015). The upper incisors have pale yellow-orange, enamel bands, and are moderately procumbent, the cheek teeth have occlusal morphology, and the toothrows are subparallel to weakly convergent. The permanent fourth upper premolar is only slightly larger than the first molar in some specimens, but P4 is conspicuously larger than M 1 in others (Voss and da Silva 2001). The dental formula is i 1/1, c 0/0, p 1/1, m 3/3, total 20, and the dentition is specialized for an herbivorous diet. Tail is short (<190 mm), average ratio of tail length to head–body length is about 50% ( Voss 2011, 2015).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Coendou vestitus Thomas, 1899
| Torres-Martínez, María M, Aya-Cuero, Carlos A, Noguera-Urbano, Elkin A, Passos, Fernando C & Ramírez-Chaves, Héctor E 2021 |
Coendu vestitus Rodríguez-Mahecha, Alberico, Trujillo, and Jorgenson, 2006:37
| RODRiIGUEZ-MAHECHA, J. V. & M. ALBERICO & F. TRUJILLO & J. JORGENSON 2006: 37 |
Sphiggurus vestitus: Honacki, Kinman, and Koeppl, 1982:572
| HONACKI, J. H. & K. E. KINMAN & J. W. KOEPPL 1982: 572 |
Coendou (Sphiggurus) vestitus vestitus:
| CABRERA, A. 1961: 603 |
Coendou (Sphiggurus) vestitus:
| TATE, G. H. H. 1935: 307 |
Coendu
| TROUESSART, E. - L. 1904: 514 |
Coendou vestitus
| THOMAS, O. 1899: 284 |