
Corallium tricolor ( Johnson, 1898 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3926.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:749A87A9-9C4E-4936-BEA9-8F99A29BEA00 |
|
DOI |
https://doi.org/10.5281/zenodo.5619921 |
|
persistent identifier |
https://treatment.plazi.org/id/932A0144-FFFD-FFE8-DEF0-9DD48E5C0029 |
|
treatment provided by |
Plazi |
|
scientific name |
Corallium tricolor ( Johnson, 1898 ) |
| status |
|
Corallium tricolor ( Johnson, 1898) View in CoL
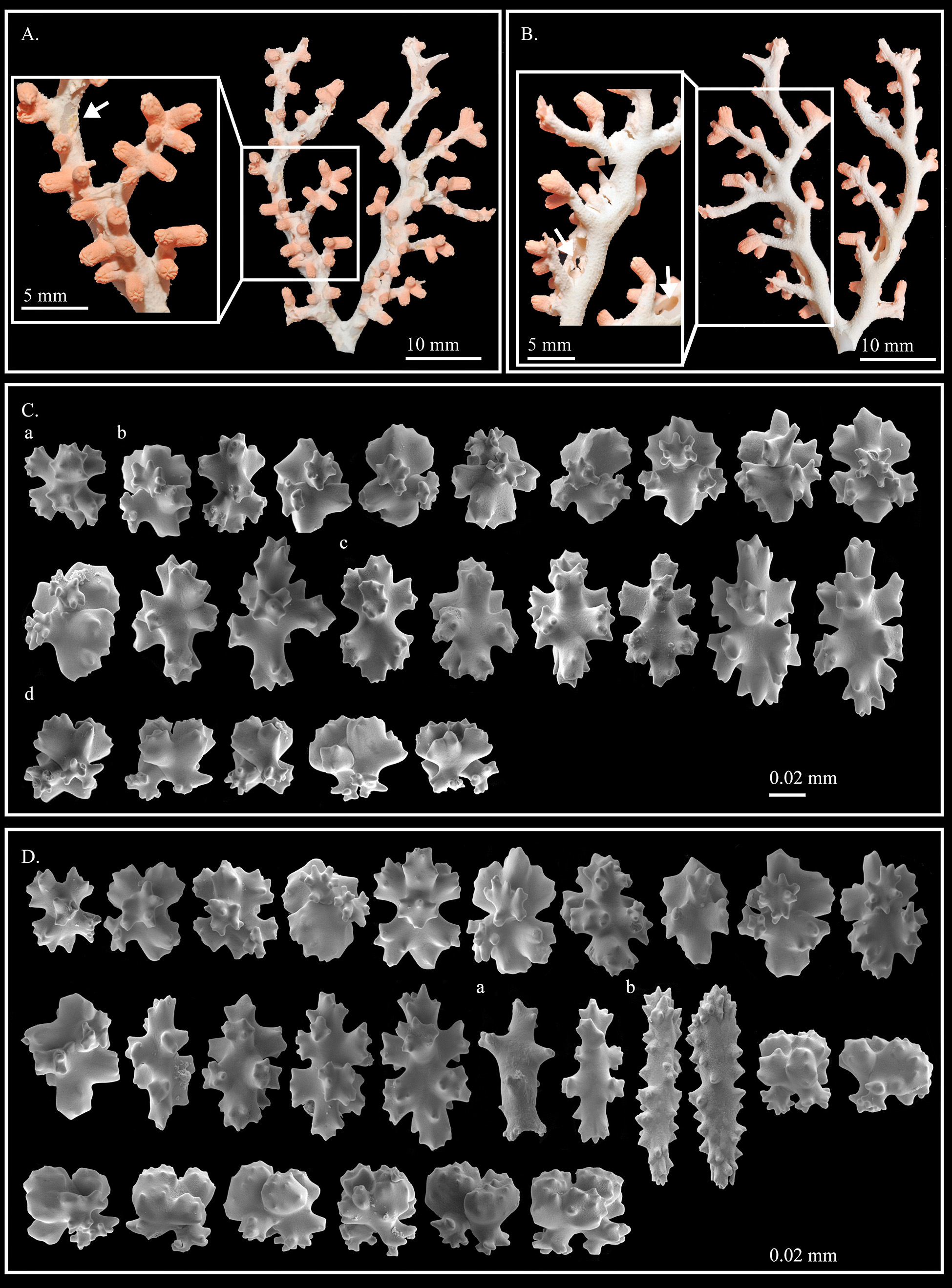
( Figure 10 View FIGURE 10 , table 2)
Hemicorallium johnsoni: Gray 1867: 126 View in CoL (partim).
Pleurocorallium tricolor Johnson, 1898: 421 .— Johnson 1899: 58, pl. 7, fig. 3.
Corallium tricolor: Kükenthal 1924: 52 View in CoL .— Bayer 1964: 467.— Grasshoff 1986: 14.— Bayer & Cairns 2003: 224.— Brito & Ocaña 2004: 290, pl. 50, fig. A–E.— Watling & Auster 2005: 292 (app. 1).— Sampaio et al. 2009: 75, fig. 1g.—Simpson & Watling 2011: 370.—Watling et al. 2011: 55 (tab. 2.1).
Material examined. SMF 9514 About SMF , Madeira (northeastern Atlantic), Jean Charcot cruise Stn. 19, 1966 , depth unknown.
Description. The colony examined is 42.20 mm tall and 33.23 mm wide, with branches sub-alternately in one plane up to the third degree of subdivision. The holdfast of the colony is lacking, and the main stem is short, 4.27 mm in height and 2.90 mm in diameter, with an elliptic cross-section. The whole colony is mainly composed of first–order branches. These are flexuous, taper from base to tip ending in slender points, are 0.78 mm in diameter at the ultimate branches, bifurcating along their length up to the third-order of branches ( Fig. 10 View FIGURE 10 A). Branchlets arising from all first, second, and third-order branches do not anastomose at the distal ends. The second and the third-order branches are nearly circular in cross-section and range in diameter from 1.34 mm to 1.75 mm. Due to the small size of this colony, the third-order branches range from 12.36 mm to 15.00 mm in length. Except for a missing holdfast, SMF 9514 is a complete specimen without fractured distal ends.
The cortex is ca. 0.057 mm thick, and it evenly wraps the axis of the colony. Close examination of the cortex reveals small raised bumps nearly uniformly covering the surface of both the autozooid-covered face ( Fig. 10 View FIGURE 10 A, indicated by an arrow), and the autozooid-free side of the colony. Due to erosion by commensal polychaetes, the axis of both first-order branches has been modified to form a cannular, tunnel-like structure composed of a thin, calcareous wall with an overlying layer of cortex ( Fig. 10 View FIGURE 10 B, indicated by a white arrow). The surface of the axis is smooth, without longitudinal grooves and is covered with minute tubercles ornamented with thorny projections.
When contracted, the autozooids are subconical in shape and almost perpendicular to the branches of colony. They have eight deep longitudinal grooves that extend from the contracted polyp orifice to halfway down the body of the polyp ( Fig. 10 View FIGURE 10 A, B). In their contracted state, autozooids measure up to 2.43 mm in height and 1.74 mm wide at their base. Autozooids are absent on the stem and are irregularly scattered on one side of the colony. At the tip of branchlets, 2–3 autozooids occur together forming a “T”. Siphonozooids are small, ca. 0.35 mm in diameter, with an obvious orifice, and surround the base of the autozooids ( Fig. 10 View FIGURE 10 B, indicated by black arrow).
Sclerome ( Fig. 10 View FIGURE 10 C, D): Cortex: there are five different types of sclerites, namely crosses, asymmetrical 6- radiates ( Fig. 10 View FIGURE 10 Ca), asymmetrical 8-radiates ( Fig. 10 View FIGURE 10 Cb), symmetrical 8-radiates ( Fig. 10 View FIGURE 10 Cc), and double clubs ( Fig. 10 View FIGURE 10 Cd). The two forms of 8-radiates are the predominant types, totally accounting for 61% of sclerites. Modified 8-radiates that resemble double clubs, with hypertrophic processes and reduced multilobular rays, account for 30% of sclerites. Some sclerites have a shape between asymmetrical 8-radiates and double clubs. Sclerites vary in size from up to 0.055 mm wide for double clubs, up to 0.059 mm long for crosses, up to 0.052 mm for 6-radiates, and up to 0.066 mm for asymmetrical 8-radiates. Symmetrical 8-radiates are the largest sclerites ranging from 0.064 mm to 0.100 mm in length.
Autozooid: polyp wall sclerites do not significantly differ from those present in the cortex. Pharyngeal sclerites are rods up to 0.081 mm in length ( Fig. 10 View FIGURE 10 Da). Tentacle sclerites include the above mentioned radiates and long rods up to 0.115 mm in length ( Fig. 10 View FIGURE 10 Db).
The colony has a pale yellow cortex with vermilion-red autozooids. The axis is uniformly lightly pink to white. Sclerites are colourless to lightly pink when examined with a microscope under transmitted light.
Distribution. Corallium tricolor is a rarely recorded species mainly distributed in the northeastern Atlantic Ocean. It has been reported from Morocco, the Canary Islands, the Cape Verde Islands, Madeira, and the Irving Seamount slope at 598–1090 m depth ( Johnson 1898, 1899; Grasshoff 1986; Brito & Ocaña 2004; Sampaio et al. 2009).
Remarks. Our description of this species is mainly based on Johnson (1898, 1899) with additions from the material examined herein. A colony collected from Madeira and deposited in the Liverpool Free Museum, was depicted by Gray (1867, p. 126) and misidentified as a young colony of Hemicorallium johnsoni . The same figure was later shown by Johnson (1899, p. 58), but it was assigned to a different species, C. tricolor . The specimen had prominent autozooids that were different from the hemispherical cortical mounds that are characteristic of C. johnsoni .
The symmetry of C. tricolor colonies varies, and the degree of branching may be related to their size. The specimen examined in this study only exhibits third-order branching, which is less than the fourth-order branching from the holotype. If compared with the holotype ( Johnson 1899, Plate VII, fig. 3), sample SMF 9514 examined herein is smaller. In the holotype, and in most autozooids of SMF 9514, the eight deep longitudinal grooves extend only to the middle of the polyp wall, but in some cases they extend to the base. In the descriptions of Johnson (1898, p. 421; 1899, p. 59), crosses were mentioned only in the autozooids, but, although scarce, they were found in both cortex and autozooids in the sample SMF 9514.
Authorship of this species is attributed to Johnson (1899). However, an incontestable valid diagnosis of the species was given before by the same author ( Johnson 1898).
Corallium tricolor is one of the species hosting the commensal polychaete Gorgoniapolynoe caeciliae (see Hartmann-Schröder 1985). The axis of the specimen studied herein has modifications induced by this worm, although the worm was lacking.
“ Coralliopsis Perrieri (Mar.)” was mentioned but poorly illustrated by Filhol (1885, p. 263, pl. 1) from a station south of the Canary Islands at 500–600 m depth. This specimen was collected during the Talisman expedition, probably in the 1883 survey. The authorship of this species is attributed to Marion (“Mar.”); however, we have been unable to find an original description or further clarifying data. A few remarks on the species were given by Perrier (1891), who considered it a synonym of C. johnsoni , as did Kükenthal (1924). In a brief discussion of this specimen, the former author provided a drawing of “ Pleurocorallium johnsoni ” collected from Cape Verde Islands by the Talisman (fig. 28, p. 73). This figure shows, however, cylindrical polyps instead of the typical low cortical mounds of C. johnsoni , and the specimen is described as having a very thin cortex, white axis, and light-red polyps.
Grasshoff (1986) only identified C. niobe and C. tricolor when he studied the material collected during the Travailleur and Talisman expeditions. The material of C. tricolor he studied was collected from the Cape Verde Islands, from where C. johnsoni was taken in abundance by the expeditions according to Perrier (1891: 72). For these reasons, although neither station from the expeditions corresponds to the coordinates given by Filhol (1885), we believe that “ C. Perrieri ” could not be a colony of C. johnsoni as has been suggested by previous authors. It is more likely C. tricolor , which was first described some years later by Johnson (1898).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Octocorallia |
|
Order |
|
|
Family |
|
|
Genus |
Corallium tricolor ( Johnson, 1898 )
| Tu, Tzu-Hsuan, Altuna, Álvaro & Jeng, Ming-Shiou 2015 |
Corallium tricolor: Kükenthal 1924 : 52
| Sampaio 2009: 75 |
| Watling 2005: 292 |
| Brito 2004: 290 |
| Bayer 2003: 224 |
| Grasshoff 1986: 14 |
| Bayer 1964: 467 |
| Kukenthal 1924: 52 |
Pleurocorallium tricolor
| Johnson 1899: 58 |
| Johnson 1898: 421 |
Hemicorallium johnsoni:
| Gray 1867: 126 |