
Ectonocryptopinae, Shelley & Mercurio, 2005
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1094.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:7CA3CB79-8B13-4FF3-B916-E812D580FBBC |
|
persistent identifier |
https://treatment.plazi.org/id/924A8786-BD04-FFA7-FEAA-66085B4DC845 |
|
treatment provided by |
Felipe |
|
scientific name |
Ectonocryptopinae |
| status |
subfam. nov. |
Ectonocryptopinae , new subfamily
Diagnosis. Minute, smallbodied Scolopocryptopidae (ca. 10–12 mm long) with generally yellowish coloration. Cephalic plate without paramedial sulci, with or without vague frontal sulcus; anterior margin slightly prolonged and sublinear, posterior margin overlapping anterior margin of tergite 1 (T1). Antennae normally with 17 articles. Coxosternum without sulci, tooth plates, lobes, or other armature. Forcipules not exceeding anterior margin of cephalic plate, without trochanteroprefemoral spurs, teeth, or processes, tarsungula substantially overlapping in closed position. T1 with prominent, anterior transverse suture giving rise to 2 converging paramedian sulci, either meeting and merging or not; T2–T22 with subparallel paramedian sulci. Ultimate tergite (T23) wider than long, caudal margin gently rounded. Legs 1–22 with stiff setae or bristles but without spines, one basal seta apiece on inner surfaces of claws in position of accessory spines. Coxopleura short, with acuminate apical spines, with or without pores. Ultimate legs subclavate in overall appearance, with 4 or 5 podomeres, without claws; prefemora and
femora with 3 and 2 ventral spinous processes, respectively, and additional small spines; tibiae and 1 st tarsi inflated and bulbous, latter with excavations on ventromedial or dorsal surfaces; 2 nd tarsi present or absent, inflated and bulbous when present.
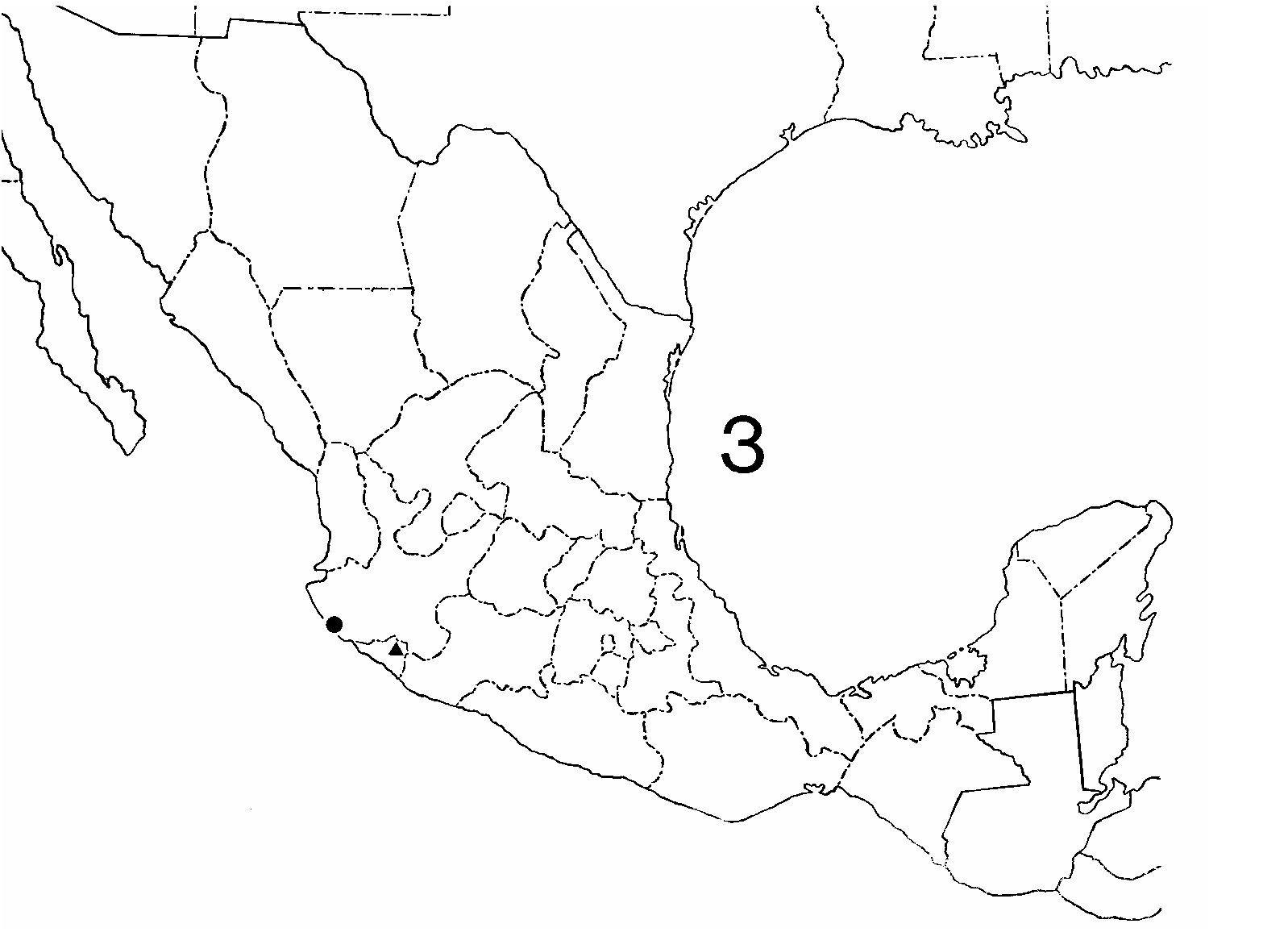
Distribution. Known only from Colima and Jalisco, Mexico ( Fig. 3 View FIGURE 3 ). The type locality of Ectonocryptops kraepelini in Colima, 3.2 km (2.0 mi) south of Tonila and around 75 km (46.8 mi) inland from the Pacific Ocean, is on the base of Colima Volcano in the Sierra Madre del Sur. Crabill (1977) gave no further details and did not report the habitat, but from internet map sites, the elevation is roughly 1,615 m (5,300 ft), and the locality is approximately 160 km (100 mi) due east of the type locality of Ectonocryptoides quadrimeropus , which is nearly at sea level (roughly 30m [98 ft]). The elevational disparity mandates substantially different habitat requirements because of markedly different environments at these heights. As the sites are in different physiographic regions, we suspect that the Ectonocryptopinae occurs widely, and in varying environments, in the western periphery of "mainland" Mexico (excluding the Baja California peninsula); it should be expected at intervening elevations and possibly even higher than Tonila. We also note that the taxon has only been encountered twice, in two Mexican states and physiographic provinces, and both samples constituted new genera and species. This suggests that field investigations from Sinaloa southward in the poorly sampled coastal border may yield more new species and possibly even genera.
Components. Ectonocryptops Crabill, 1977 ; Ectonocryptoides , new genus.
Relationships. We present in fig. 4 a hypothesis of subfamilial relationships in the Scolopocryptopidae ; outgroups are the two other scolopendromorph families, Scolopendridae and Cryptopidae . Apomorphies are as follows:
1. Presence of 23 pairs of legs and pedal segments. This is the apomorphy for the Scolopocryptopidae ; the only other scolopendromorph with this feature is the Brazilian scolopendrid, Scolopendropsis bahiensis (Brandt, 1841) ( Kraepelin 1903, Attems 1930, Bücherl 1974, Schileyko 2002). All other scolopendromorphs possess 21 leg pairs and segments.
2. Presence of at least one ventral "spinous process" on each ultimate prefemur. Lewis et al. (2005) suggested the term "spinous processes" for the projections from the ultimate legs in the Scolopocryptopidae , which are immovable outgrowths of the exoskeleton, are not connected to it by articulations, and do not arise from sockets or alveoli. These structures are absent from the Kethopinae but present in, and apomorphic for, the clade, Scolopocryptopinae + ( Ectonocryptopinae + Newportiinae ). They differ from true "spines" found in the Scolopendridae , which can be dislodged leaving a faint, rounded hump.
3. Presence of at least one ventral "spinous process" on each ultimate femur. In addition to those on the prefemora, the Ectonocryptopinae and Newportiinae exhibit "spinous processes" on the ventral surfaces of the ultimate femora, which constitutes a synapomorphy for these subfamilies.
4. Loss of claws. To our knowledge, claws are present on the ultimate legs of all scolopendromorphs except representatives of the Ectonocryptopinae and Newportiinae . This loss is another synapomorphy for these subfamilies and is possibly associated with the assumption of a sensory function that manifests itself differently in the taxa. The elongated, "pseudoarticulated" caudal legs of newportiines superficially resemble antennae and would seem to have a similar function, while the inflated, subclavate appendages of ectonocryptopines possess apical depressions with fine protruding hairs that are consistent with sensory structures, although their presence in Ectonocryptops awaits confirmation.
5. Presence of a dorsomedial "spinous process" on each ultimate prefemur. Both components of the Scolopocryptopinae possess an additional, smaller "spinous process" on the dorsomedial surface of each ultimate prefemur; Chagas (2003) reported that this structure is on the dorsolateral surface of D. miersii , but it is actually dorsomedial (Chagas, pers. comm. to RMS). The Ectonocryptopinae and Newportiinae possess true "spinous processes" only ventrad, so the presence of a small dorsomedial one is a synapomorphy for Dinocryptops + Scolopocryptops and an autapomorphy for the Scolopocryptopinae .
6. Presence of a trochanteroprefemoral spur. All scolopocryptopines possess this autapomorphic structure; while varying in size, it is at most only a large spur and never a longer process as in the Scolopendridae . It is absent from the other subfamilies except for minute spurs in a few species of Newportia .
7. Inflated, bulbous podomeres. To our knowledge, such podomeres, which impart a subclavate overall appearance to the ultimate legs of ectonocryptopines and are soft and weakly sclerotized, are not found on any other scolopendromorph. The enlarged caudal podomeres in representatives of the Plutoniuminae (Cryptopidae) and some scolopendrids are quite different, being stiff, rigid, and strongly sclerotized ( Shelley 1997).
8. Small size. The minuteness of presumably adult ectonocryptopines, ca. 11 mm (0.4 in.) long and less than 1.0 mm wide, contrasts them with all other scolopocryptopids, which are considerably larger, well over 25 mm (1.0 in.) long as adults.
9. Pseudoarticulations. In the Scolopendromorpha , division of the ultimate 2 nd tarsi into "pseudosegments" is unique to the Newportiinae .
No apomorphy is known for the Kethopinae , which we regard as the plesiomorphic sistergroup to the rest of the Scolopocryptopidae . As noted by Crabill (1960), it is distinguished by flexure of the ultimate 2 nd tarsi and the presence of "saw teeth" on the tibiae and 1 st tarsi, these being plesiomorphies that are shared with the Cryptopinae ( Cryptopidae ) (see Crabill 1958, fig. 3; 1960, fig. 15) and seemingly indicative of shared ancestry. The Kethopinae is endemic to, and occurs sporadically in, the aforementioned western and southwestern United States ( US), its present stronghold being the western slope of the Sierra Nevada in California, where Kethops utahensis (Chamberlin, 1909) has been encountered most often ( Shelley 2002). Additionally, Thalkethops grallatrix Crabill, 1960 , seems rather common in the caves of Carlsbad Caverns National Park, this being a form that apparently sought refuge from the arid desert environments of southeastern New Mexico in cool, moist subterranean habitats ( Crabill 1960, Barr & Reddell 1967, Shelley 2002). Despite several field trips to epigean areas where the Kethopinae occurs, RMS has never seen a live individual, and preserved ones also are rare, as none exist in California repositories. We believe the Kethopinae is an "old" group that is declining naturally toward extinction, and it may have been displaced from much of its former range by the younger and more successful Scolopocryptopinae , which is at or near its zenith, at maximal expansion with minimal geographic discontinuities. A Pangean origin for both subfamilies is likely and would account for the latter's occurrences in Africa and Asia.
The Scolopocryptopinae surely antedates the Newportiinae and Ectonocryptopinae , with more regional distributions. Crabill (1977) correctly perceived that Ectonocryptops View in CoL is related to Newportia View in CoL , but the affinity is actually at the subfamilial level. These taxa appear to have arisen comparatively recently and have "experimented" with modifications of the distal podomeres of the caudal legs for a possible sensory function, an aspect of which is loss of the claw. In Ectonocryptoides quadrimeropus View in CoL , the proximal podomeres are heavily armored with small spines in addition to the "spinous processes"; Crabill (1977) does not mention such spines in Ectonocryptops kraepelini View in CoL and perhaps did not examine the appendages at high enough magnification, a matter to be checked on fresh material. The Newportiinae is quite successful, having dispersed across the Panamanian portal and spread over most of the Neotropics including the West Indies; the subfamily therefore contrasts with the entire order Geophilomorpha View in CoL , which shows little evidence of trans Panamanian passage ( Pereira et al. 1997). The present restricted distribution of the Ectonocryptopinae is surely an artifact, though the taxon probably is not as widespread as the Newportiinae .
The Scolopocryptopinae , Ectonocryptopinae , and Newportiinae occur in Mexico, while the Kethopinae lies just to the north in the adjacent US, a geography that suggests a broad part of the now southwestern US and northern/northwestern Mexico as the likely source area for the family. Consequently, if undiscovered scolopocryptopid "subfamilies" exist, this region, particularly the Sierra Madre, is where they likely occur.
Remarks. As no specimen or illustration exists, we necessarily have to accept Crabill's accounts (1977) for features of Ectonocryptops kraepelini in formulating the foregoing diagnosis. It may thus need modification when this species or the genus is rediscovered, and the accuracy of Crabill's written descriptions can be assessed. He did not, for example, characterize the apices of the caudal legs, so we do not know whether they also exhibit depressions with protruding hairs; if so, this will be another apomorphy of the subfamily.
According to Crabill (1977), the holotype of Ectonocryptops kraepelini had 17 articles in both antennae. One individual of Ectonocryptoides quadrimaculatus also has 17, but the other has 2 & 3 fewer on the left and right antennae, respectively. This disparity is slight, and we believe 17, the basic number of antennomeres in the Scolopendromorpha , to be the number in the Ectonocryptopinae .
The apparent rarity of the Ectonocryptopinae probably reflects a general lack of investigations along the Pacific Coast of Mexico coupled with their minuteness and a possible preference for litter habitats. A person collecting manually would have difficulty distinguishing an organism this small in litter or a mass of decaying leaves, whereas the centipede would stand out somewhat by contrast under bark of decaying logs, as do species of Cryptops Leach, 1815 (Cryptopidae) in the eastern United States. Consequently, Berlese extraction is probably the best, and perhaps the only, method of collecting ectonocryptopines. An analogous situation in Mexico involves the rarely encountered milliped order Siphoniulida , which was known only from Sumatra and Guatemala until seven individuals were discovered in berlesate samples from Chiapas and Veracruz ( Sierwald et al. 2003). The authors surmised that siphoniulids may be deep humus dwellers that are missed by investigators focusing on larger, subsurface forms and concluded that "berlese extraction may be the best method of obtaining siphoniulids, and one should probably include substantial humus as well as loose litter."
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Ectonocryptopinae
| Shelley, Rowland M. & Mercurio, Randy 2005 |
Ectonocryptopinae
| Shelley & Mercurio 2005 |
Ectonocryptoides quadrimeropus
| Shelley & Mercurio 2005 |
Ectonocryptops kraepelini
| Shelley & Mercurio 2005 |
Ectonocryptopinae
| Shelley & Mercurio 2005 |
Ectonocryptopinae
| Shelley & Mercurio 2005 |
Kethopinae
| Shelley 2002 |
Ectonocryptops
| Crabill 1977 |
Scolopocryptopinae
| Pocock 1896 |
Newportiinae
| Pocock 1896 |
Newportiinae
| Pocock 1896 |
Newportiinae
| Pocock 1896 |
Scolopocryptopinae
| Pocock 1896 |
Newportiinae
| Pocock 1896 |
Newportia
| Gervais 1847 |