
Krenakanthus ribeiranus Leme, Gonella & D.R. Couto, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.619.1.2 |
|
DOI |
https://doi.org/10.5281/zenodo.8425691 |
|
persistent identifier |
https://treatment.plazi.org/id/9175B60B-2829-FFCC-FF15-FBA9FC1AEA4F |
|
treatment provided by |
Plazi |
|
scientific name |
Krenakanthus ribeiranus Leme, Gonella & D.R. Couto |
| status |
sp. nov. |
Krenakanthus ribeiranus Leme, Gonella & D.R. Couto , sp. nov. ( figs. 1–11 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 )
Diagnosis:—This new species is morphologically related to K. roseolilacinus , but distinctly differs by stemless habit (vs. long caulescent), leaf blades subdensely to densely covered by spreading, hair-like trichomes (vs. glabrescent or glabrous), margins straight (vs. undulate mainly toward the base), inconspicuously spinulose and appearing entire (vs. distinctly spinulose), inflorescence shortly pedunculate to subsessile (vs. sessile), simple (vs. sparsely branched only at the base), flowers distinctly smaller (ca. 30 mm vs. 42–55 mm long), sepals smaller (11–11.5 × 2.5 mm vs. 20–27 × 4–5 mm), petals smaller (25–26 × 8 mm vs. 35–44 × 15–20 mm), and exappendiculate (vs. bearing distinct cupuliform appendages).
Type:— BRAZIL. Minas Gerais: Alvarenga, Serra de Santa Maria , maciço quartzítico a leste da sede do município, [precise locality withheld for conservation purposes], 650 m elevation, 1 March 2023, P.M. Gonella 3760, J.C.S. Ribeiro, D.P. Cordeiro, L.D. Martins & J. Martins (holotype RB!, isotypes BHCB!, CESJ!, MBML!) .
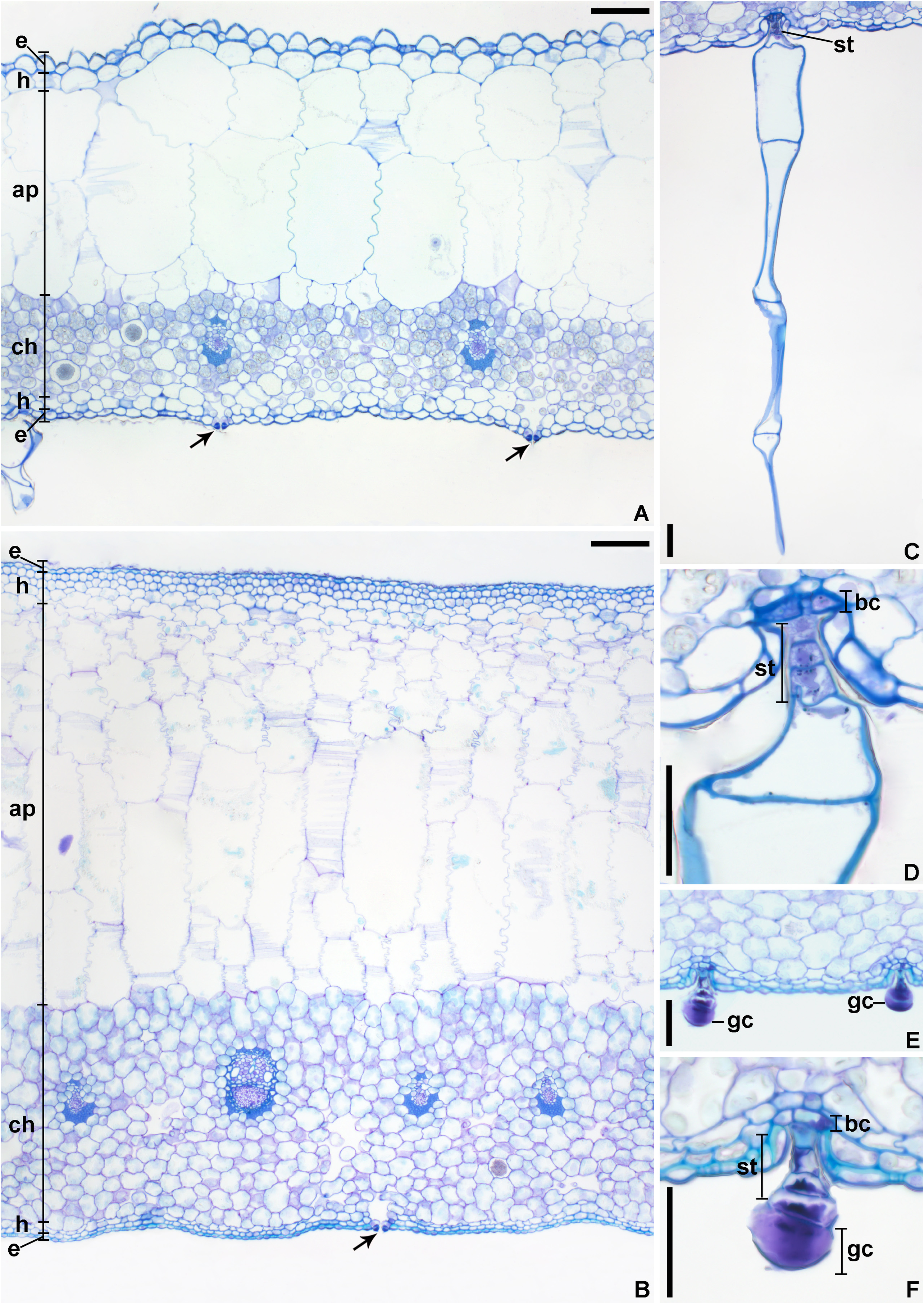
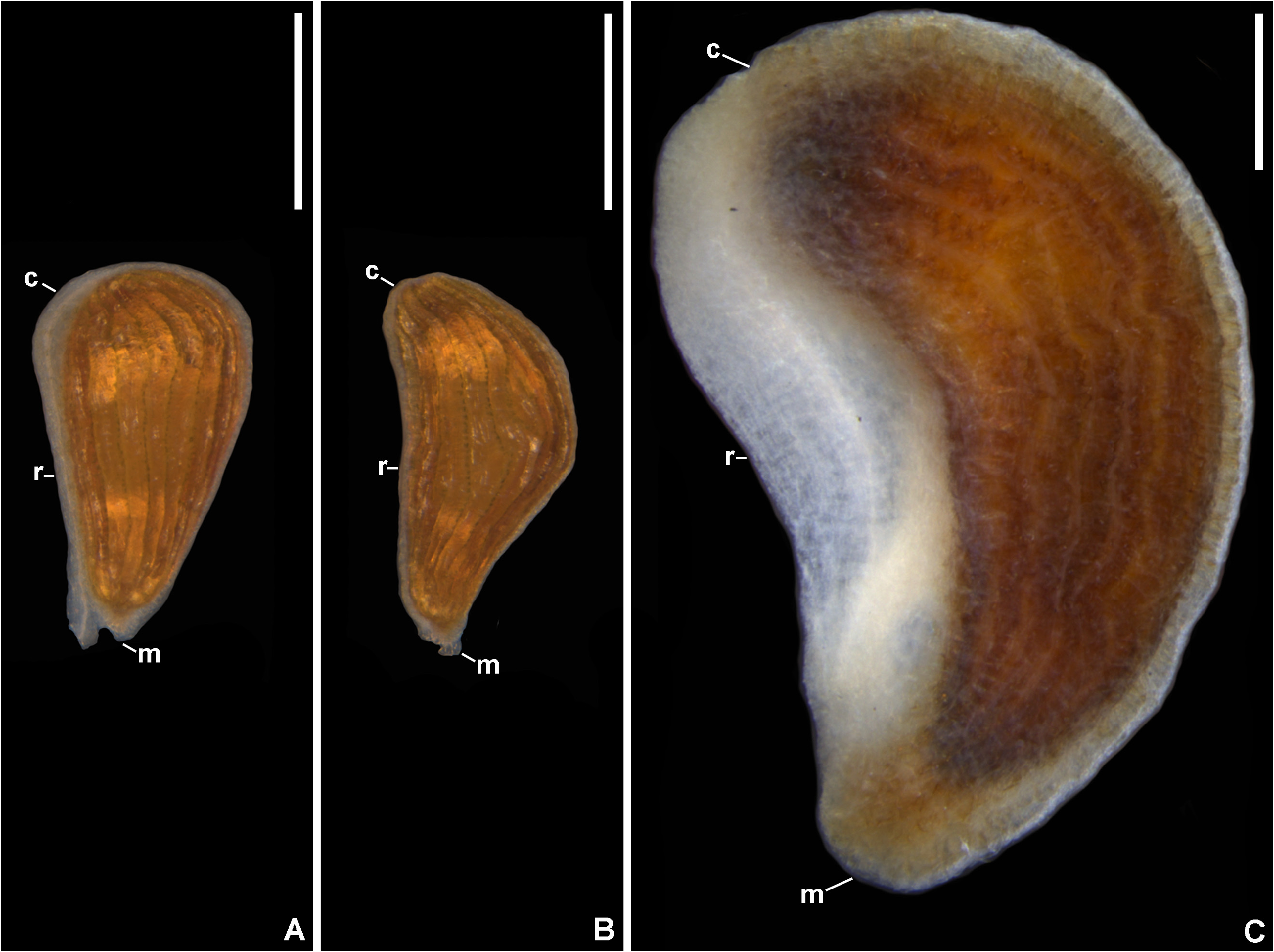
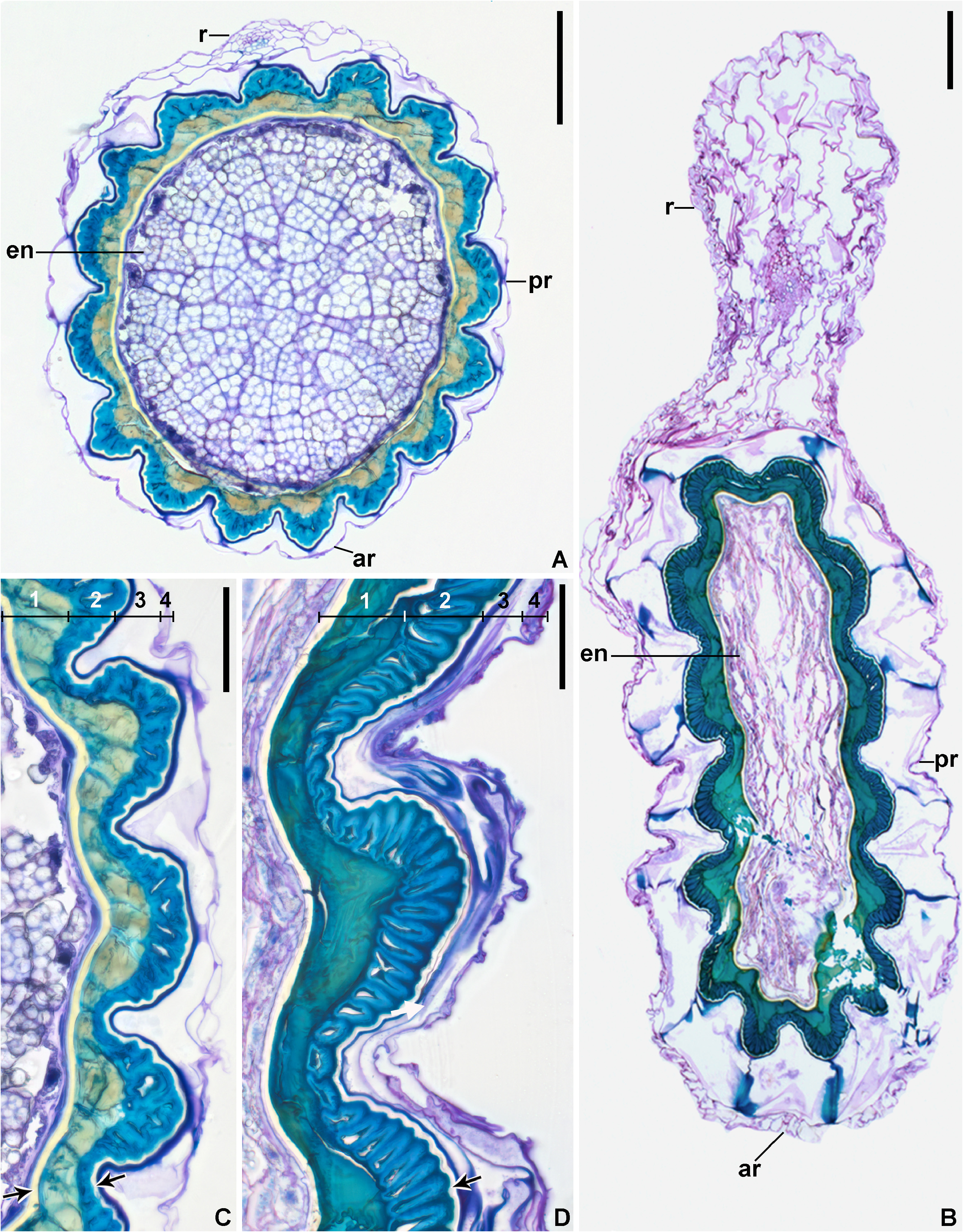
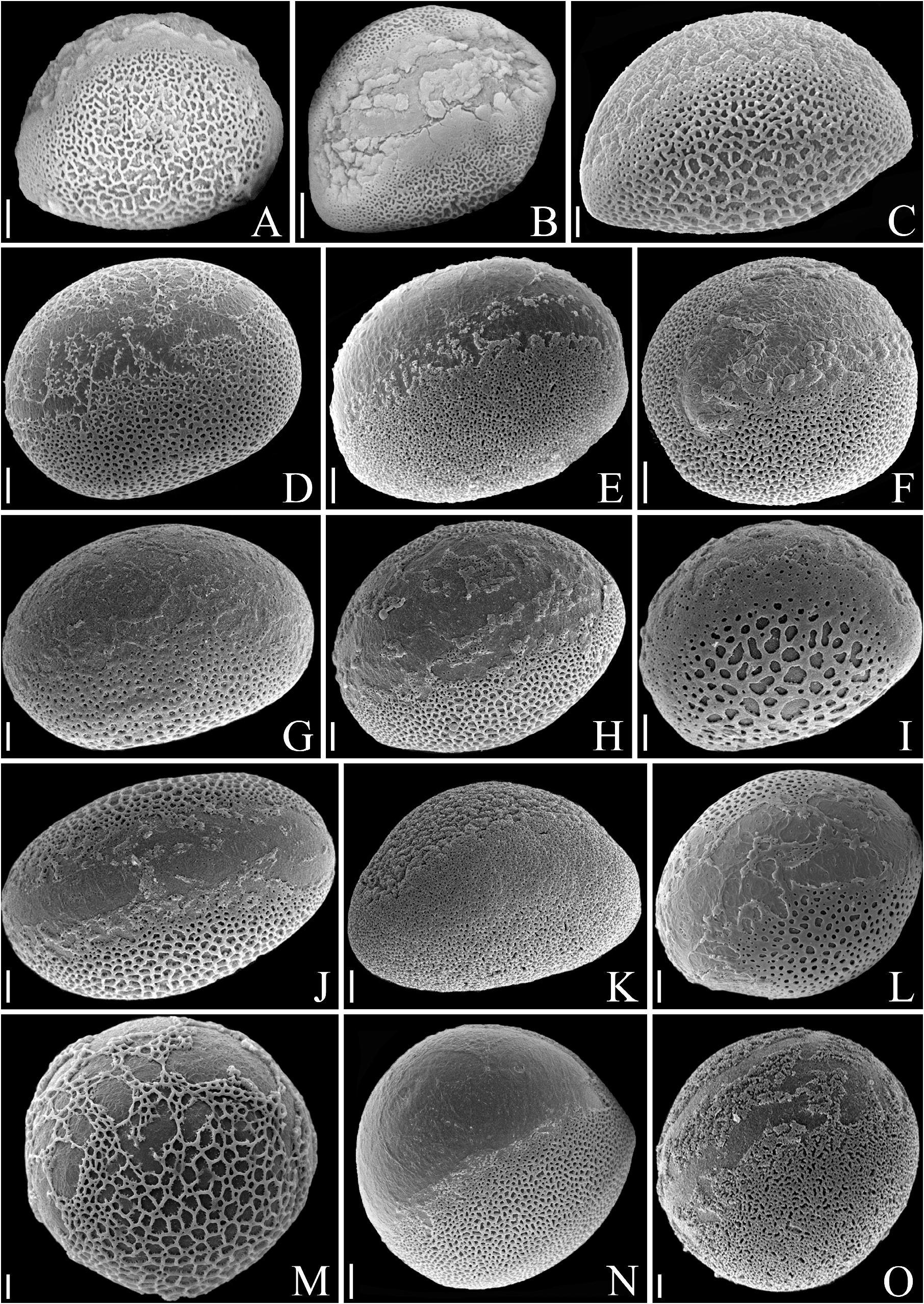
Description:— Plants saxicolous, stemless, flowering 2.5–3 cm heigh, propagating by short basal shoots. Leaves 7–12 in number, very thin in texture, spreading, prostrate, forming a lax rosette without water impounding capacity; sheaths inconspicuous, ca. 9 × 7 mm, remotely spinulose, nerved, with sparse, long hair-like trichomes; blades narrowly lanceolate, narrowed toward the base, apex acuminate-caudate, 7–25 × 0.8–1.8 cm, green, concolorous, opaque, finely nerved, subdensely to densely covered by spreading, uniseriate hair-like trichomes mainly abaxially, margins straight, with dense long hair-like trichomes, inconspicuously spinulose and appearing entire; spine s whitish, narrowly triangular, 0.1–0.2 mm long, 1–2 mm apart, spreading (upper ones) to slightly antrorse (basal ones). Inflorescence shortly pedunculate to subessile, 2–2.5 cm long (not including the petals), ca. 1.5 cm in diameter (not including the petals), simple, corymbose; peduncle 5–8 mm long, ca. 2 mm in diameter, greenish-white, with spreading uniseriate hair-like trichomes; peduncle bracts foliaceous; floral bracts the basal ones foliaceous, distinctly exceeding the flowers, the upper ones lanceolate, acuminate-caudate, very thin in texture, green, spreading-recurved, shorter than the sepals, with spreading uniseriate hair-like trichomes, finely nerved, ecarinate, 5–10 × 2–3.5 mm, margins entire. Flowers 5–10 in number, all perfect, sessile, ca. 30 mm long (with the petals extended), fragrance not detected; sepals 11–11.5 × 2.5 mm, lanceolate, acuminate, entire, connate at the base for 4–4.5 mm, green but drying dark castaneous soon after anthesis, with sparse spreading hair-like trichomes, thin in texture, the adaxial ones carinate with the keels continuing on the ovary, the abaxial one obtusely if at all carinate; petals broadly spathulate from a very narrow base, 25–26 × 8 mm, 3.1–3.2 times longer than wide, free, green at the base and lilac toward the apex, the blades broadly elliptic to broadly obovate, acute, spreading at anthesis and forming a fan blade-like corolla, flaccidescent afterwards, at the base with 2 longitudinal callosities equaling the antepetalous filaments, with sparse and inconspicuous glandular trichomes, exappendiculate; stamens deeply included and not visible; filaments distinctly unequal in length, the antepetalous ones ca. 3.5 mm long, adnate to the petals for ca. 0.5 mm, the antesepalous ones ca. 5 mm long, free, slightly complanate, greenish-white toward the distal end; anthers 1.2–2 mm long, oblong, dorsifixed at ca. 1/4 from the base, base bilobed, apex distinctly apiculate; pollen oblate, 25–35 μm in diameter, sulcate, sulcus narrow, with exine elements, margins weakly defined to indistinct, exine reticulate, reticulum broadly meshed proximally and meshes decreasing in size towards the sulcus margins; style about equaling the antepetalous stamens and shorter than the antesepalous ones, white; stigma conduplicate-spiral, whitish, blades elongate, papillose, erect; ovary 3–4 mm long, ca. 3 mm in diameter distally, trigonous, white, with sparse hair-like trichomes; epigynous tube lacking; ovules numerous, obtuse; placentation axial, median to apical. Fruits subglobose, greenish becoming pale yellowish castaneous to castaneous, 4 × 3.5–5 mm, with persistent sepals, 2.7–2.9 times longer than the fruit length; seeds at least 90 in number, obovoid to subtrapeziform, 0.8–1.2 × 0.6 mm, yellow, distinctly longitudinally sulcate.
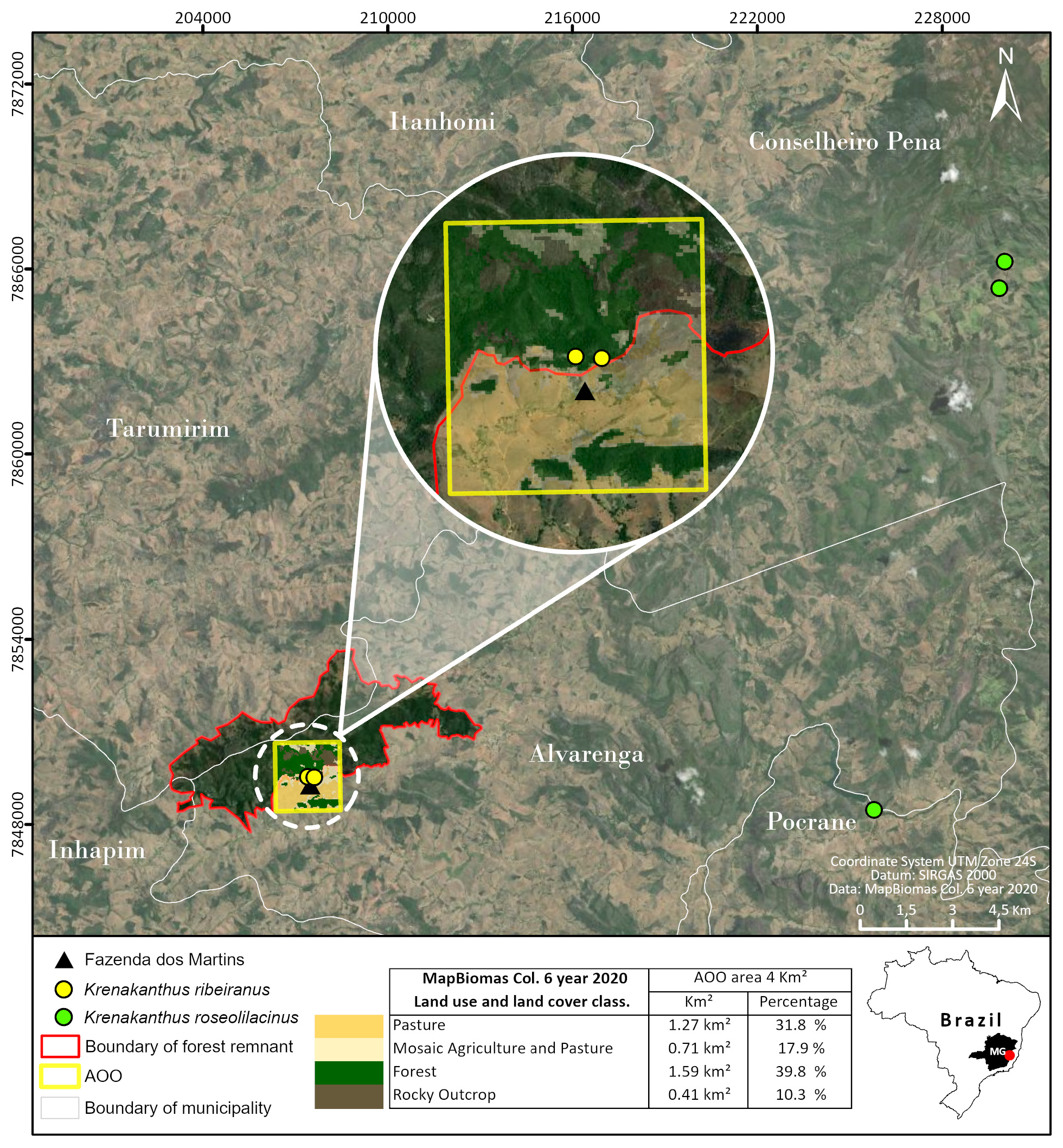
Distribution and habitat:— Krenakanthus ribeiranus is a sciadophytic, saxicolous species known from two small subpopulations located in the type region, the Serra de Santa Maria, Alvarenga county, Minas Gerais state ( figs. 1–2 View FIGURE 1 View FIGURE 2 ). The species habitat is a riparian forest in a matrix of Semideciduous Seasonal Forest and Campos Rupestres in the Atlantic Forest domain, where it is found in a small fragment of an advanced regeneration of secondary forest.
The only known two subpopulations of the species are located very close to the forest edges, which directly adjoins pasture areas. The first subpopulation, located near a waterfall, contains approximately 200 mature individuals and a large number of juveniles. The species appears to have a high recruitment rate, with many germinating seeds and young plants taking root ( fig. 5 F View FIGURE 5 ). Seed germination occurs in close proximity with mother plants, and occasionally within the fruit, indicating limited dispersal capacity, which may explain the restricted range of the species.
The second subpopulation contains approximately 100 mature individuals, located on a rock wall by the river about 10–15 m from a pasture. This subpopulation is particularly vulnerable to fire events that could seriously impact the species, not only directly but also by altering the deeply shaded environment that is critical for its survival, given its high shade demand.
Both subpopulations are located at about 550–650 m elevation and are composed of scattered individuals growing on organic-rich, shallow soils accumulated on sandstone rock surfaces among mosses, in shaded rocky spots alongside creeks ( fig. 3 View FIGURE 3 ).
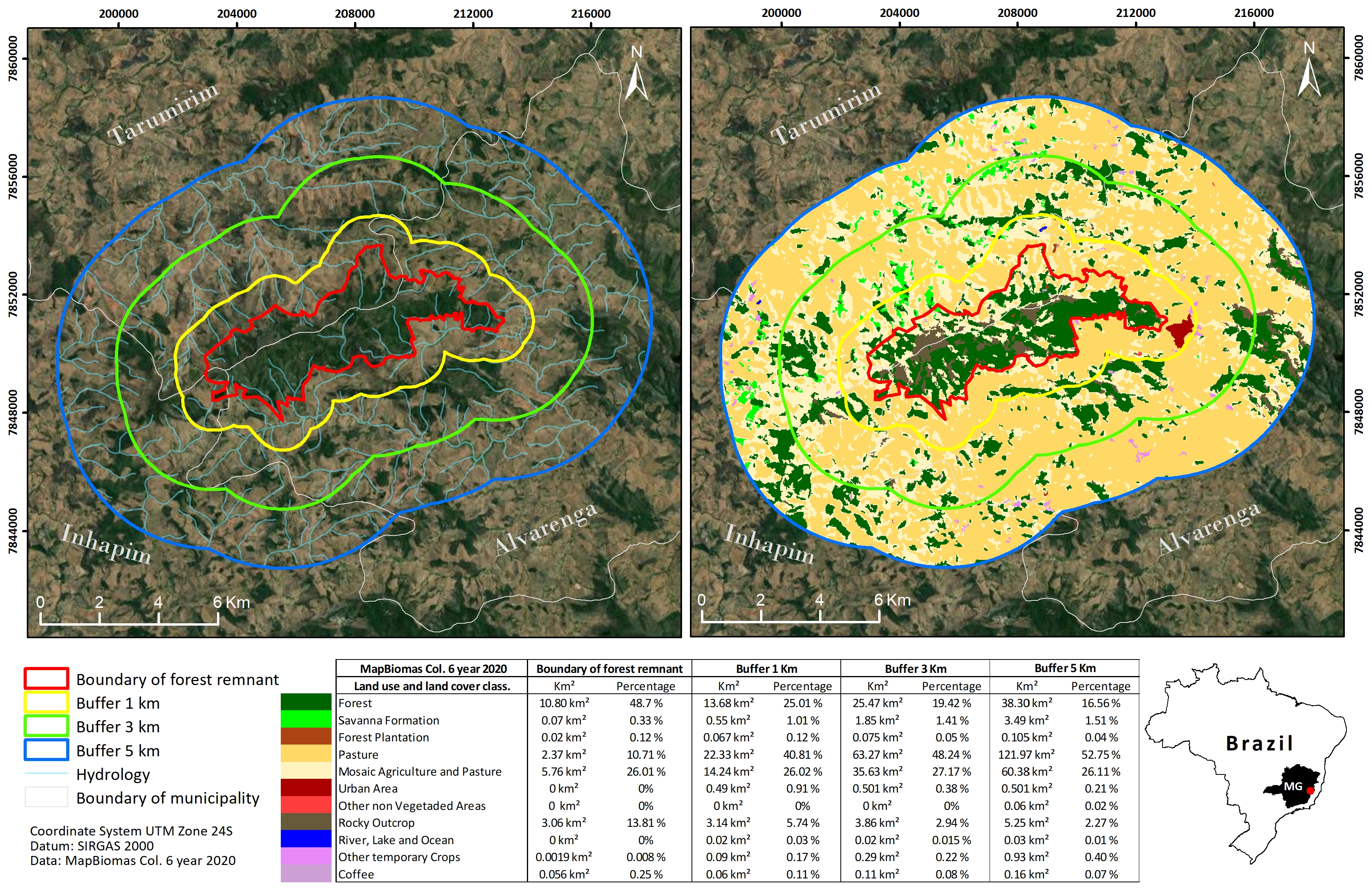
The Serra de Santa Maria range is situated on the boundary of Alvarenga, Inhapim, and Tarumirim counties. Despite sharing a similar geomorphological history with other ranges in the area, such as Pico da Aliança in Alvarenga county, it is geographically isolated and predominantly composed of quartzitic Campos Rupestres and semideciduous forest remnants. The range is inadequately protected by any designated protected area and has been poorly sampled botanically. Human activities have resulted in severe habitat conversion, exacerbating the range’s biota vulnerability.
The applied overlay analyses (Jord„o et al. 2022) showed that the massif is suffering from intense human-induced disturbances and that only a small fraction of the range is suitable for the species ( fig. 2 View FIGURE 2 ). Although 48.7% of the total massif area is still forested, and 13.81% is rocky outcrops, we have only located K. ribeiranus near small creeks under gallery forests, with a much smaller actually occupied area than the total AOO and the area of the range. This makes it highly vulnerable to the direct and indirect effects of agriculture and pasture expansion, which comprise nearly 26% of the total massif area. Additionally, 10.71% of the massif is utilized for temporary crops, which uses fire to clear the land and eventually promote intense fire events. Even in this scenario, however, the massif is a strong candidate for the creation of a private or public conservation unit, as the region’s deficit in terms of in situ conservation strategies suggests.
Etymology:—The epithet of this new species honors its discoverer, the young naturalist Júlio Cesar dos Santos Ribeiro, that lives at the foothills of Serra do Padre Ângelo, Conselheiro Pena, Minas Gerais State. With his sharp eye, escalating skills, and great curiosity for the natural world, Mr. Ribeiro has dedicated himself intensively to the field survey of the species that make up the extremely rich flora of his homeland, having participated in the discovery of many of the novelties from the region, such as K. ribeiranus .
Conservation status:—Critically Endangered—CR B1ab (ii,iii,v) + B2ab (ii,iii,v). Several threats affect both subpopulations, such as agriculture (coffee plantations and other temporary crops), deforestation due to cattle ranching and grazing, fires, which already affected one subpopulation, invasion by alien grasses ( Urochloa sp. , used in the surrounding grazing areas), which can synergistically intensify fire events. Although targeted surveys in other areas of similar habitat may potentially reveal unrecorded subpopulations, so far, the species can be considered a narrow endemic bromeliad from the last significantly continuously forested massif of this region.
Since the species is currently known only from two nearby collection points or subpopulations, it does not have an associated EOO polygon and has an estimated AOO of 4 km ². Land use and land cover data retrieved from Mapbiomas (2022a, b) applied for the overlay analyses (Jord„o et al. 2022) indicates that over 31% of its AOO was converted to pastures, and that 17.9% is currently designated as a mosaic of agriculture and pastureland formations ( fig 1 View FIGURE 1 , 2 View FIGURE 2 ). Considering the intensity of the described stress vectors, one single location is considered, as these threats may extirpate the whole population if one single intense stress event occurs within its narrow and fragile habitat. Although we have an indication of the putative number of mature individuals for each known subpopulation, no data on generation length and the population trend is known, so the species could not be assessed based on its (a priori small) population size and reduction rate. All known subpopulations are outside protected areas, which might increase the vulnerability of the species in face of severe anthropogenic events.
Therefore, based on the minimum values of EOO and AOO, combined with one single location and an estimated continuing decline in its AOO, extent and quality of habitat, and possibly in the number of mature individuals, the species is here declared as Critically Endangered under the aforementioned IUCN (2012) criteria.
Additional specimens examined(paraypes):— BRAZIL. Minas Gerais: Alvarenga, Serra de Santa Maria, Sobreiro de Cima , maciço quartizítico a leste da sede do Município, [precise locality withheld for conservation purposes], 595 m elevation, 27 November 2022, J.C.S. Ribeiro 001, cult. E. Leme 10220 ( RB!); ibidem, 615 m elevation, 1 March 2023, P.M. Gonella 3800, J.C.S. Ribeiro & D.P. Cordeiro ( RB!) .
Distinctive characters:—The set of morphological features of this new species makes its circumscription in Krenakanthus apparently challenging. This is true due to the several morphological differences when compared to the single known species of the genus, K. roseolilacinus . These differences are: (a) stemless habit ( figs. 3 C–D View FIGURE 3 , 5 B–C, E View FIGURE 5 ; vs. long caulescent) and much fewer leaves (7–12 vs. 20–27 in number); (b) leaf blades subdensely to densely covered by spreading, hair-like trichomes ( figs. 4 C–D View FIGURE 4 , 7 A View FIGURE 7 ; vs. glabrescent, with minute glandular trichomes, or glabrous, fig. 4 A–B View FIGURE 4 ), smaller (7–25 × 0.8–1.8 cm vs. 17–27 × 1.8–2.4 cm), thinner in texture, margins straight ( fig. 5 A–B View FIGURE 5 ; vs. undulate mainly toward the base), inconspicuously spinulose and appearing entire ( fig. 4 C View FIGURE 4 , 7 B View FIGURE 7 ; vs. distinctly spinulose, fig. 4 A–B View FIGURE 4 ), spines inconspicuous [0.1–0.2 mm vs. 0.3 (apical ones)– 3 mm (basal ones) long]; (c) inflorescence shortly pedunculate to subsessile ( fig. 7 C–D View FIGURE 7 ; vs. sessile), simple (vs. sparsely branched only at the base and simples toward the apex); (d) flowers distinctly smaller (ca. 30 mm vs. 42–55 mm long); sepals smaller (11–11.5 × 2.5 mm vs. 20–27 × 4–5 mm), shorter connate at the base for 4–4.5 mm ( fig. 6 G View FIGURE 6 ; vs. 3–7 mm, fig. 6 H View FIGURE 6 ); (e) petals smaller (25–26 × 8 mm vs. 35–44 × 15–20 mm), apex acute ( figs. 5 D View FIGURE 5 , 6 A, C View FIGURE 6 ; vs. acute to rounded, fig. 6 B, E View FIGURE 6 ), and exappendiculate (vs. bearing distinct cupuliform appendages); (f) fruits smaller, 4 × 3.5–5 mm (vs. 8–10 × 7–12 mm); (g) seeds smaller, 0.8–1.2 × 0.6 mm (vs. 2–3 × 0.7–1 mm), costal bands 8–12 cells wide, intercostal bands 1–3 cells wide (vs. costal bands 15–20 cells wide, intercostal bands 4–6 cells wide).
Despite the relevant differences that distinguish these two species of Krenakanthus from each other, the similarities that unite K. ribeiranus and K. roseolilacinus are: (a) leaves thin to very thin in texture; (b) peduncle inconspicuous to absent); (c) sepals distinctly connate at the base; (d) petals similarly free, broadly spathulate from a very narrow blade ( fig. 6 C View FIGURE 6 ), blades spreading at anthesis and forming a fan blade-like corolla, and flaccidescent afterwards ( fig. 6 A View FIGURE 6 ); (e) stamens deeply included and not visible at anthesis ( fig. 6 A–B View FIGURE 6 ); (f) filaments distinctly unequal in length ( fig. 6 C–D View FIGURE 6 ); (g) stigma conduplicate-spiral ( fig. 6 M–N View FIGURE 6 ), being the only two representatives of the “Cryptanthoid complex” with such a stigma type; (h) epigynous tube lacking; and (i) fruits with persistent sepals 2.5–2.9 times longer than the fruit length ( fig. 6 I–J View FIGURE 6 ). In addition, both species present similar habitats, occurring in the understory of Semideciduous Seasonal Forest in the region of Alvarenga and Conselheiro Pena ( fig. 1 View FIGURE 1 ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.