
Acanthocyclops americanus (Marsh, 1893)
|
publication ID |
https://doi.org/ 10.1080/00222933.2012.744432 |
|
persistent identifier |
https://treatment.plazi.org/id/8F6C9A6D-FFE6-7E43-E8EF-FF0BFCA9600D |
|
treatment provided by |
Felipe |
|
scientific name |
Acanthocyclops americanus (Marsh, 1893) |
| status |
|
Acanthocyclops americanus (Marsh, 1893)
Acanthocyclops robustus f. limnetica Petkovski, 1975.
Acanthocyclops robustus Kiefer (1976) part., Fryer (1985), Dodson (1994), Lescher- Moutoué (1996), Einsle (1996).
Acanthocyclops trajani Mirabdullayev and Defaye 2002 .
Material examined. All populations of this species used in genetic analyses are registered in Table 1, and include material from americanus terra typica localities (previously mentioned pond in Madison, Wisconsin, leg. Dr S. Dodson and a pond in city park, Washington DC, USA, leg. Dr V. Ivanenko) and the A. trajani type locality (Etang de Noes, vicinity of Paris, coll. V. Alekseev).
Additional material. USA: pond in California, leg. Dr B. Kuperman; Canada: pond in Waterloo, Ontario, leg. Dr H. Fernando; Canada: Lake Pin-Rouge, Montreal University field station, Quebec, 9 October 1998, coll. V. Alekseev; Mexico: pond in Mexico City, 12 October 2009 coll. V. Alekseev; Russia: small river in the River Volga delta, May 1984, coll. V. Alekseev; Russia: small lake in northeast Tchukotka, June 1985, leg. Dr P. Krylov; Russia: the River Ob delta, western Syberia , leg, Dr V. Kuzikova; Russia: Posolsky Bay , Baikal, central Siberia , 2 July 1986, coll. V. Alekseev; Russia: rice fields, Krasnodar vicinity, July 1987, leg, O. Feraponova; Russia: lake in tundra, Tchaun, Magadan vicinity, 15 July 1982, leg, Dr E. Streletzkaya; Russia: pond in Pushkin Town , Leningrad district, 28 June 1997, coll. V. Alekseev; Latvia: Riga Bay, 22 July 1997, coll. V. Alekseev; Armenia: Lake Sevan , 4 July 1990, coll. V. Alekseev; Ukraine: pond in Krymia peninsula, Simferopol vicinity, 2 May 1990, coll. V. Alekseev; Ukraine: pond in Kiev vicinity, August 2011, coll. V. Monchenko; Norway: pond in City Park, Oslo, 23 June 2009, coll. V. Alekseev; Germany: canal in Oldenburg, July 1997, coll. V. Alekseev; Belgium: pond in university botanical garden, Ghent, 23 June 1989, coll. V. Alekseev; France: pond in botanical garden, Boulogne-sur-Mer, 20 July 2007, coll. V. Alekseev; Spain: Ebro River delta, June 2003, coll. M. R. Miracle; Spain: Guadalquivir River , Sevilla, 4 January 2009, coll. V. Alekseev; Egypt: River Nile , Cairo, 12 April 2003, coll. V. Alekseev .
Redescription of the neotype female and allotype male. Female neotype dissected on slide N 56736 and placed into the type collection of Zoological Institute of the Russian Academy of Sciences, St Petersburg, Russia. The type series also includes two females and one male preserved in pure glycerol .
Female neotype. Body transparent, colourless. Full body length without furcal seta 1217 µm. Egg sacs with 35– 40 eggs each. Cephalosome as long as wide, with maximum width close to its end ( Figure 3A View Figure 3 ).
Genital double somite 1.1 times as long as wide, with seminal receptacle with wide transparent zone in its frontal part ( Figures 3A–B View Figure 3 ). Anal segment with row of small dense dents, proctodeum with single row of setules on both sides. Furcal rami parallel without hairs on inner margin, five times as long as wide. Length proportions of distal setae, beginning from outer terminal seta: 1.0/5.3/9.0/2.1. Inner seta sub-equal to furca length. Dorsal seta about 1.2 times as long as inner setae and lateral setae about half length of outer setae.
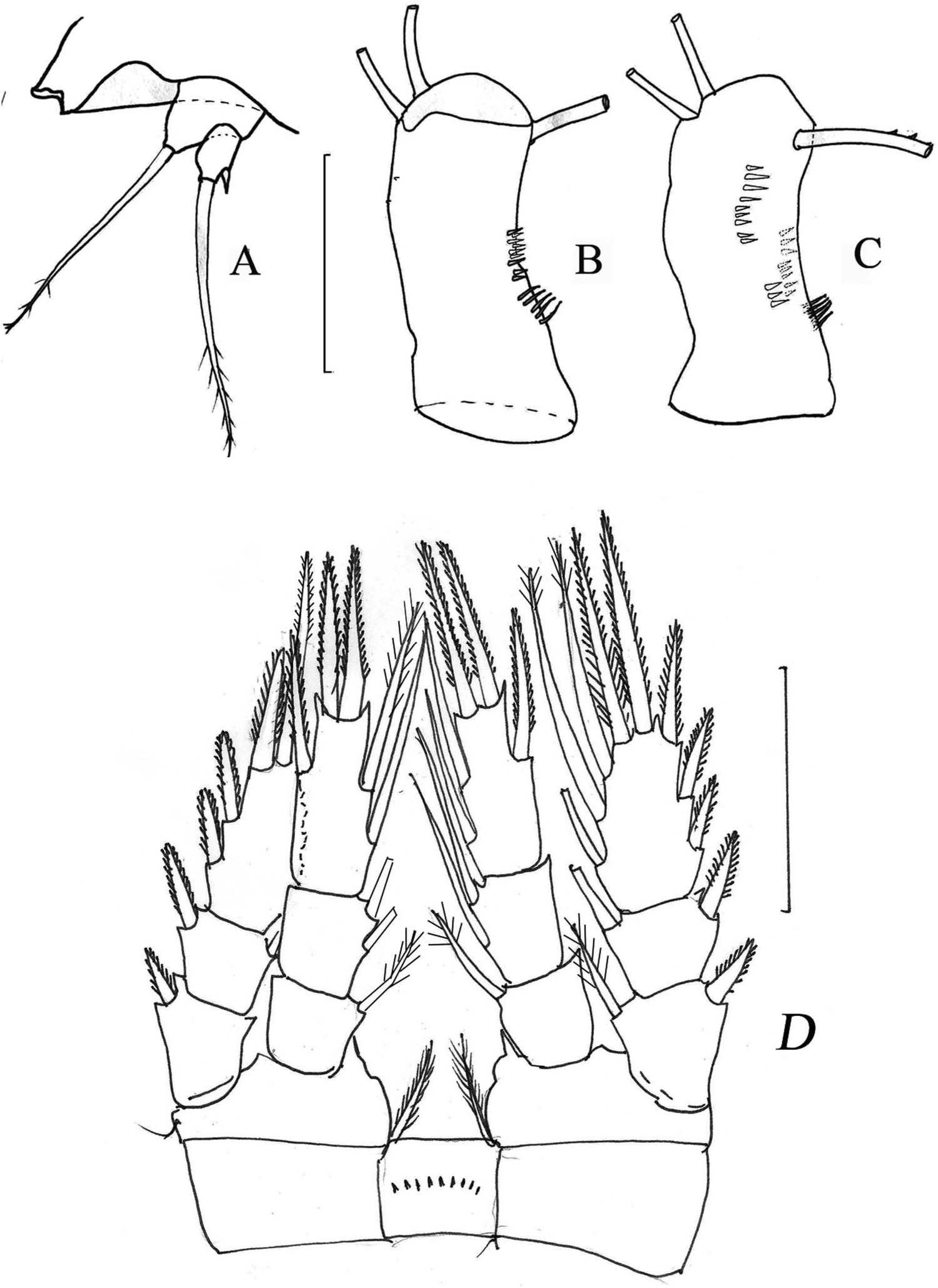
Antennule of 17 segments, not reaching distal margin of first thoracic somite. Setation of antennular segments (aesthetascs in Roman numbers) beginning from first: 8/4/2/6/3+I/1/2/1/1/0/1+I/2/0/1/2/2+I/7+I. Shortest setae of distal segment less than length of distal segment ( Figure 4A View Figure 4 ). Antennal basipodite at caudal side with three lateral groups of hair-setae in middle ( Figures 4B View Figure 4 and 6B View Figure 6 ). At frontal side this segment with two groups (9+3) of strong denticules on top, long line of 15–17 relatively small spinules in the central part and two groups of several spinules as shown in Figure 6C View Figure 6 . Third segment bearing nine seta and last segment bearing seven seta.
Gnathobase of mandible with six teeth, rudiment of endopodial segment with two long setae and short seta ( Figures 4C View Figure 4 ). Maxillula with three strong and three small teeth, two strong setae; maxillar palp with seven setae, different in length ( Figure 4D View Figure 4 ). Maxilla of five segments, praecoxa with two strong setae in its middle part; coxa with a strong seta in the middle and an endite bearing two claw-like setae; basal endite with two very strong claw-like spines, both with a row of spinules and small setae near the place of fusion of the rudimentary endopod. Endopodite 1 with three clawlike setae and endopodite 2 bearing distally two long setae ( Figure 4E View Figure 4 ). Maxilliped of four segments, praecoxa + coxa with two strong setae in the middle part and small setae at their distal end; basis with two setae of different length and three groups of strong spinules near the insertion of the setae; first segment of endopod with strong spine and rudimentary endopod, bearing a strong spine and two hairless setae ( Figure 4F View Figure 4 ).
Swimming legs 1–4 with three-segmented rami ( Figure 5A–D View Figure 5 ). Distal segments of exopod of legs 1/2/3/4 with 3/4/4/4 spines, respectively. Distal segments of endopod of Legs 1–2 with one spine and five setae. Distal segment of endopod Legs 3–4 elongated, with two strong spines at its end. In Leg 4 this segment 2.6 times as long as wide and inner apical spine 1.1 times as long as outer spine. Insertion of the lateral outer seta/spine approximately at the same distance from the end of the segment as the second lateral inner seta. Intercoxal sclerite Leg 1 with high hills, and without hairs on its free edge. Intercoxal sclerites Legs 2–3 also with small hills on free edge and without hairs on its free edge ( Figure 5A–C View Figure 5 ). Intercoxal sclerite Leg 4 with small hills and row of small denticules in middle part. Coxa Leg 4 with four groups of dents typical for several species of robustus group ( Figure 5D View Figure 5 ). Rudimentary Leg 5 two-segmented, basal segment with long outer seta. Distal segment with long apical seta and short inner spine less than half of segment length ( Figure 6A View Figure 6 ).
Male allotype. Dissected and mounted in glycerol surrounded with Canadian balsam on slide N 56737. One male from the type series is preserved in pure glycerol (N) and placed in slide covered with cover glass with plasticine tombs and not compressed. All material is in the collection of the Zoological Institute of the Russian Academy of Sciences (Reg. N 56736–56738). Body length without furcal seta 1030 µm.
Cephalosome 1.3 times as long as wide, with maximal width close to end of its length ( Figure 3C View Figure 3 ). Caudal rami 4.3 times as long as wide, slender inner seta more than twice length of outer spine-like seta. Lateral seta placed without spinules in its base. Dorsal seta placed near inner seta insertion, about 1.2 times as long as outer seta.
Antennule 12-segmented. Setation of antennular segments (aesthetascs in Roman numbers) beginning from first: 7( III)/3/1/6(I)/1/1/1/4(I)/0/1/0/7. Antennal basopodite ornamentation as in female. Morphology of mouth appendages and Leg 1–3 basically as in female.
Distal segment of endopod Leg 4 about three times as long as wide, with inner apical spine about as long as the segment and 1.25 times as long as outer apical spine ( Figure 6D View Figure 6 ). Insertion of lateral seta/spine in this segment as in female. Inner edge of basis of Leg 4 with short hair-setae, coxa of Leg 4 with strong spine. Coxa Leg 4 on its caudal side, with same groups of dents as in female but fewer in number. Intercoxal sclerite Leg 4 without hills and hair-setae on its free edge, but with short row of small spinules in middle. Rudimentary Leg 5 two-segmented, with setae and spine of similar proportion to female. Rudimentary Leg 6 with inner spine, middle seta slightly shorter than spine and a very long outer seta (relative lengths 1.0/0.75/2.1) that clearly separates it from other species of this group.
Ecology. Planktonic, often dominating in eutrophic water bodies.
Distribution. Described from North America, it is widely distributed in North and Central America, in the tropics mainly in mountain lakes. In Europe it was first found in the beginning of the twentieth century in England, but since then it has been frequently found in all European countries (named A. robustus , after Kiefer’s revision). Its area seems to be expanding occurring mainly in coastal water bodies, fish ponds and reservoirs. It has been found also in Africa, Asia and South America.
Phenotypic plasticity. Swimming legs spine formula is seasonally variable as well as the conversion to spine of the lateral seta of endopodite 3P4 (this gave rise to the name A. americanus spinosus Monchenco, 1961 ).
Collections from the Natural Park of Albufera of Valencia have been reanalyzed to look for phenotypic variability. This Natural Park is a coastal wetland near the town of Valencia, consisting mainly of a coastal lagoon surrounded by a marshland now transformed into rice fields and irrigation channels where the dominant species is A. americanus , previously referred to as A. robustus ( Oltra and Miracle 1992; Alfonso and Miracle 1990). As in other localities, A. americanus in Albufera of Valencia showed a significant variability in size: females, 850–1300 µm and males, 670–990 µm, according to summer–winter development. Spine formula of exopods was mostly 3444/3444, and only a small fraction of individuals showed some variation. In summer a few individuals had the formula 3443/3443 and in winter we found individuals with 3444/ 4444, although other combinations were also possible, always with an important degree of asymmetry, such as 3344/3444. Dodson (1994) described a similar morphological seasonality in his “ A. robustus ” specimens and separated them into two forms, “cold-water” and “warm-water”, based on the number of spiniform versus setiform setae on the terminal segment of the Leg 4 exopod.
In Albufera of Valencia Natural Park there are also seasonal variations in the dominance of forms of the typicus and spinosus type. The seasonal spinosus character of P4 endopodite seems to be related to factors, other than temperature, affecting diapause and development. In the lagoon and channels, which are permanent water bodies, the typical form of A. americanus is the most abundant and common one and only from November to February, a small proportion of the population was A. americanus spinosus . Whereas in rice fields, which are temporary habitats, the dominant form is A. americanus spinosus present also in summer. From these results we relate the presence of the spinosus form with a population coming from diapause. The rice fields are dry during two periods: late summer–autumn and late winter–early spring, so their A. americanus population may have in summer as well as in winter individuals coming from diapause stages. In temporary intradunal ponds of this park the form spinosus is also the most frequent type found. The spine in the P4 endopodite may be of help when they move out of the sediment. In the genetic analyses we have included both forms A. americanus from Albufera of Valencia, with seta and with spine.
Molecular-genetic results
Diversity in the COI gene
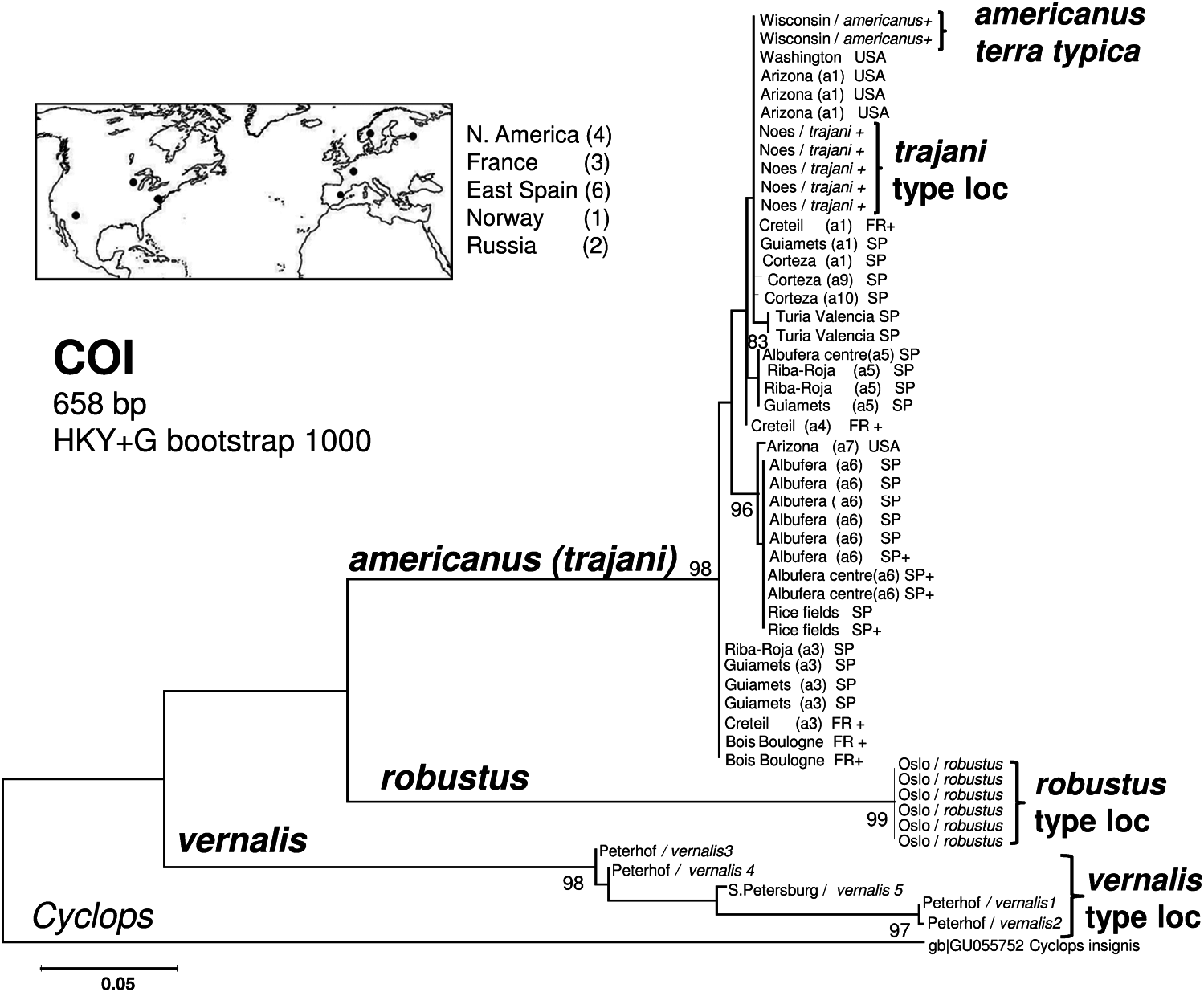
In total, 52 individuals from 3 species and 1 form variant ( Table 1) were sequenced for 658 nucleotides giving 15 haplotypes, which contained 460 variable sites and 139 parsimony informative sites. All phylogenetic methods (ML, Bayesian and parsimony) resulted in the same topology. The ML topology is presented in Figure 7 View Figure 7 . Our data
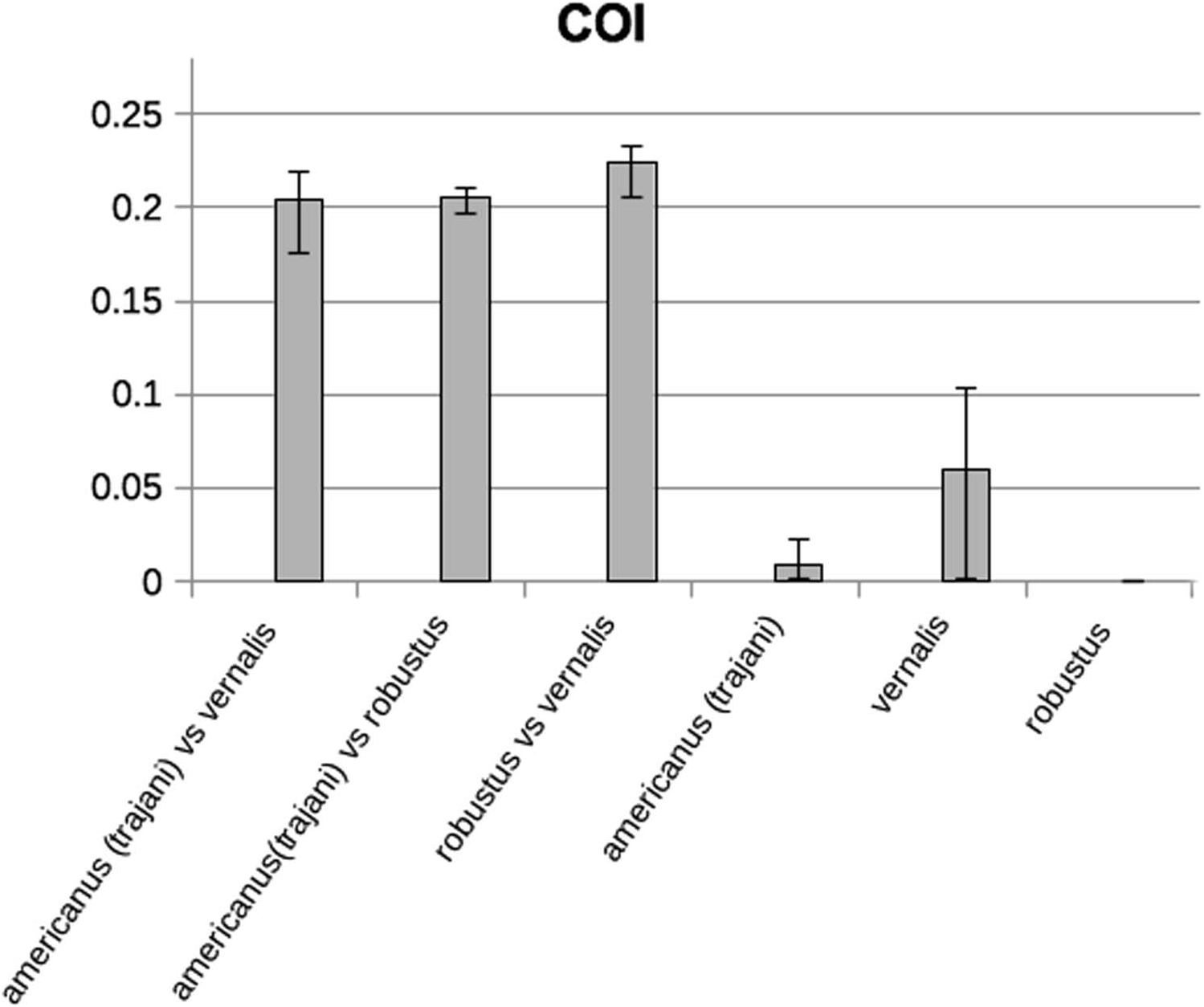
show three clearly differentiated well-supported lineages, corresponding to the three species described at the end of the nineteenth century: robustus , vernalis and americanus . Genetic sequence divergence (K2P distance for haplotypes) between these three lineages was very high – around 20% ( Figure 8 View Figure 8 ). Using minimum–maximum pairwise K2P distances, divergence times among these species may be estimated at 10–14 My, assuming a strict molecular clock hypothesis and a substitution rate of 1.66% per My, taken from COI calibrations for Decapoda, owing to the absence of specific calibrations for the rate of substitutions in Copepoda. The general rate of COI evolution was estimated at 1.66–2.30% per My for shore-crabs by Schubart et al. (1998) and at 1.40% per My for mangrove alpheid shrimps by Knowlton and Weigt (1998).
K2P genetic distances were also calculated taking into account only non– synonymous substitutions (codons 1 and 2), giving average distances of 0.032, 0.015 and 0.029 between americanus – vernalis , americanus – robustus and robustus – vernalis , respectively. These distances are also quite high.
The six individuals of A. robustus from the type locality (two samples taken on the same day) were grouped together without differences between them as a totally separate clade. The five individuals of A. vernalis from the type locality (four samples from Peterhof, and one sample from a nearby area) also resolved as a totally separate clade, but with a relatively high diversity among the four haplotypes (up to 10%; Figure 8 View Figure 8 ). All the other 41 individuals clustered together in 1 clade, corresponding to the species A. americanus (recently renamed A. trajani ), with very small genetic divergence within them. A. americanus individuals from the type region of the first description, Wisconsin, USA, had exactly the same sequence as the individuals from Etang de Noes, France, the type locality of A. trajani . Furthermore, most specimens from other USA localities as far apart as Washington and Arizona also had this exact same sequence, together with specimens from a Spanish reservoir and Lake Creteil in France. Within one site, it is common to have two or three haplotypes which are also found at other sites. The other haplotype from the USA was almost coincident with haplotypes from the coastal lagoon Albufera of Valencia. The analyzed individuals within this clade have 11 haplotypes in total, but the maximum sequence divergence between them was 2.2% ( Figure 8 View Figure 8 ). Distances taking into account only non-synonymous substitutions within species were 0.001 for americanus and 0.007 for vernalis , confirming the homogeneity of americanus haplotypes and the higher divergence within the vernalis lineage.
The specimens of the form spinosus (inner spine in end3P4) within the americanus clade had exactly the same sequence as the individuals without this character (typical form with seta instead of spine in inner side of end3P4) from the same site. Individuals from the Albufera coastal lagoon taken on the shore and in the centre, as well as the individuals from the surrounding rice fields, had the same sequence regardless of whether they were the form spinosus or the typical form with seta.
Diversity in 12S rRNA gene
PCR-amplification efficiency was much higher for the 12S gene than for COI. Most of the individuals sequenced for COI were sequenced for 12S, but since 12S is slightly less variable than COI some redundant individuals were not used for the analysis; therefore, a total of 19 individuals were sequenced from 3 species and 1 form variant ( Table 1) for 426 sites. For this 12S gene, there were 22 existing haplotype sequences (343 sites) of this species complex in GeneBank, all derived from the work of Bláha et al. (2010) and they have been included in the phylogenetic tree for comparison. Thus, we worked with a total of 441 sequences, corresponding to 25 haplotypes that contained 107 variable and 88 parsimony informative sites. As for COI, the different phylogenetic methods produced congruent trees; all predicted the existence of four well-differentiated lineages. The topology of the ML tree is presented in Figure 9 View Figure 9 . The reconstructed phylogeny shows the same three clades corresponding to the same three species separated by COI plus a new clade represented only by GenBank haplotype sequences. This corresponds to an apparently new species of this group, not found in our samples. Genetic divergence (evaluated with K2P [ Figure 10 View Figure 10 ]) between these clades, as in COI, was very high – around 20% (ranging from 16 to 25%). The two individuals from the type locality of A. robustus were clustered together with one haplotype from GenBank resulting from individuals collected in the Czech Republic and in Slovakia and identified as A. einslei by Bláha et al. (2010). This indicates that the newly described species A. einslei is a synonym of A. robustus . The four individuals from the type locality of A. vernalis clustered together with A. vernalis haplotypes from GenBank from individuals collected in the Czech Republic and identified by Bláha et al. (2010). Two had exactly the same sequence as the most frequent GenBank haplotype found in this clade. Variability within this clade was also relatively high, up to 5.2%, but less than within the clade resulting from COI (with only our specimens from the St Petersburg area). The individuals identified as A. americanus in the present study from several localities and from the type locality of A. trajani clustered together with the so-called A. trajani from GenBank collected in several localities of the Czech Republic, two in Greece, one in Portugal and one in western Spain. Altogether, the mitochondrial gene 12S was slightly less variable than COI with six haplotypes and very limited variability between them, ranging from 0.3 to 1.9%. The haplotype from the type locality for A. trajani was identical to the USA haplotype and to one of the haplotypes from eastern Spain. The two Greek haplotypes from GenBank were identical to two eastern Spain haplotypes and one from Lake Creteil, whereas those from the Czech Republic, Portugal and western Spain from GenBank were very similar but not identical to the ones analyzed in the present study.
As found with COI, the spinosus form of A. americanus from the Albufera area has the exact same sequence as the typical form from the same area.





| V |
Royal British Columbia Museum - Herbarium |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Acanthocyclops americanus (Marsh, 1893)
| Miracle, M. R., Alekseev, V., Monchenko, V., Sentandreu, V. & Vicente, E. 2013 |
Acanthocyclops robustus Kiefer (1976)
| " sensu Kiefer 1976 |