
Curtonotum taeniatum Hendel
|
publication ID |
https://doi.org/10.11646/zootaxa.3079.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/8F1187DF-680B-FF82-FF38-F994FAD1F8E5 |
|
treatment provided by |
Felipe (2021-08-23 20:28:45, last updated by Plazi 2023-11-04 13:54:50) |
|
scientific name |
Curtonotum taeniatum Hendel |
| status |
|
Curtonotum taeniatum Hendel View in CoL
Figures 131–137 View FIGURES 131–137
Curtonotum taeniatum Hendel, 1913: 629 View in CoL .— Enderlein, 1917: 69 [records from Callanga, Peru and Songo, Bolivia].— Wirth, 1975: 78.2 [synonym of Curtonotum gibbum (Fabricius) , Neotropical catalogue]. Thompson and Pont, 1994: 80 [synonym of and replacement name for Musca gibba Fabricius ].
Musca gibba Fabricius, 1805: 297 . Preocc. Müller, 1776.
Curtonotum gibbum (Fabricius), 1805 .— Macquart, 1843: 350 [combination].— Malloch, 1930: 325 [as senior synonym of Curtonotum taeniatum Hendel View in CoL ].— Aldrich, 1905: 641 [Nearctic catalogue]. Curran, 1934: 440 [as senior synonym of Curtonotum taeniatum Hendel View in CoL , key, British Guyana list].— Wirth, 1975: 78.2 [Neotropical catalogue].
Diplocentra gibba (Fabricius), 1805 .— Giglio-Tos, 1895: Parte Quarta: 35 [combination, Mexican catalogue].
Diagnosis. Characters to distinguish this species from C. flavisetum are provided in the diagnosis of that species.
No external characters have been found to reliably distinguish C. taeniatum from C. tumidum . Curtonotum tumidim often has pale setulae on the anterior face of the fore femur dorsal of the ctenidial comb (both species have varying amounts of pale setulae proximal of the ctenidial comb), pale vibrissae, and very few dark medial setae on the mid coxae, whereas no C. taeniatum specimen examined has pale setulae dorsal of the ctenidial comb (in one specimen they are dark brown), only a few have a pale vibrissa (none have both vibrissae pale), and only one has fewer than three dark setae on each mid coxa. Too few specimens have been examined to say whether these small differences in chaetotaxy colour can reliably separate these species.
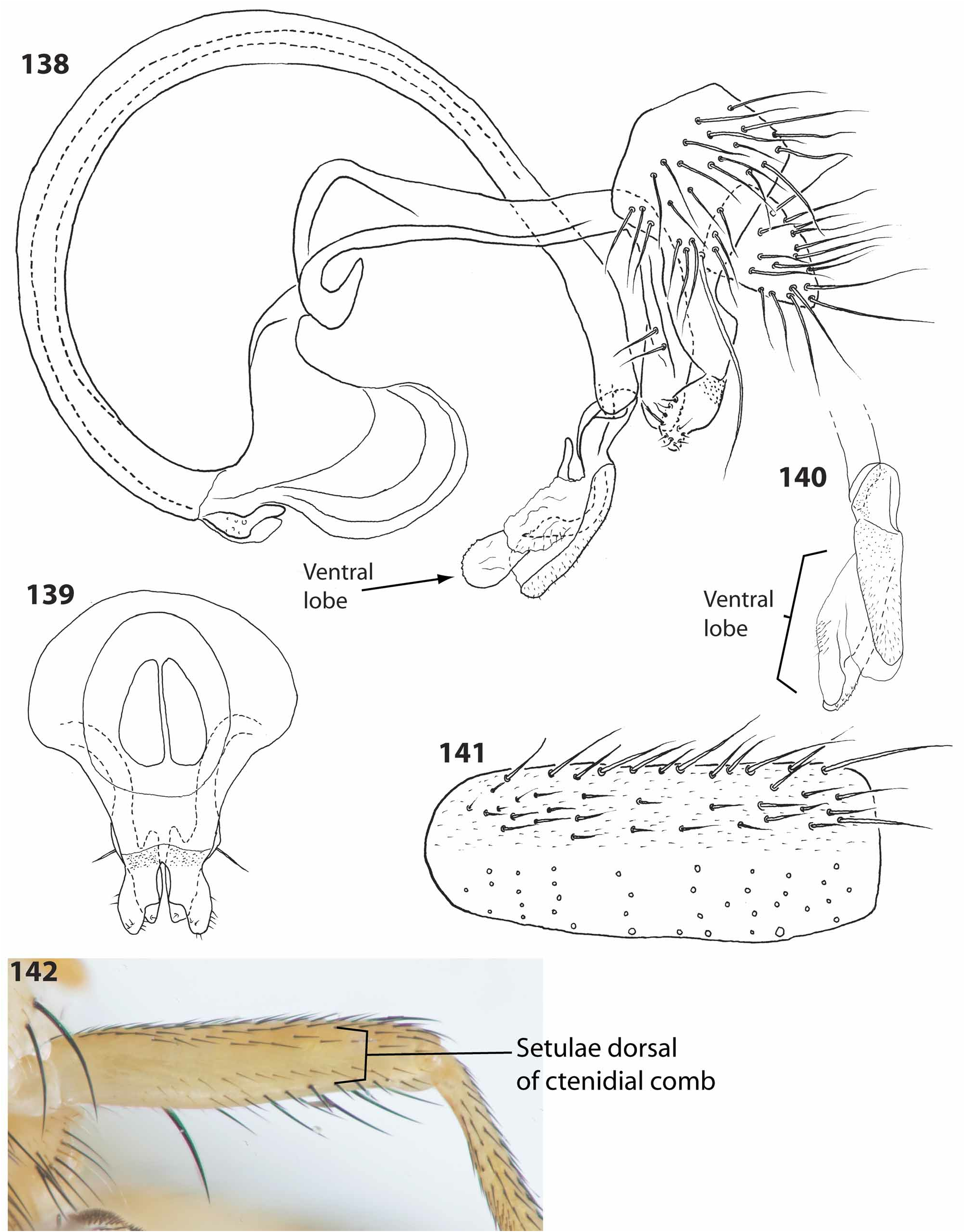
Males of C. taeniatum can be distinguished from C. tumidum by the relatively stout ventral distiphallic lobe (this lobe is very elongate and apically bent in C. tumidum ; compare Figures 131 and 133 View FIGURES 131–137 with 138 and 140). Identification of females is more problematic, as discussed in the comments section below.
Description. Similar to C. flavisetum except as follows. Length: 7.0– 9.8 mm. Frons width 1.3–1.4 times frons height. Eye height 10.3–16.7 times gena height. Vibrissa much larger than adjacent subvibrissals, all subvibrissals usually black (in some specimens a few of these are pale), two male specimens from Madre de Dios, Peru, and the two specimens from Cururipe, Bahia, Brazil, each with 1 (either the right or the left) vibrissa orange-yellow. Scutum often with pair of lateral vittae, extending from posterior of transverse suture to posterior margin. Postpronotal lobe often with additional moderately long seta anterior to 3 prominent setae. Anepisternum with 3 very large and 2–5 moderate setae on posterior half. Prominent seta at posteroventral corner of katepisternum, and usually 1–2 adjacent setae, black. Mid coxa with at least 3 dark setae medially, usually many more. Setulae on the anterior face of fore femur dorsal of the ctendial comb black or dark brown, several setulae proximal to the ctenidial comb yellow.
Male terminalia: Sternite 5 narrow, with sparse yellow and black setulae, posterior margin truncate and well sclerotized (as in Figure 141 View FIGURES 138–142 ); tergite 6 poorly sclerotized, desclerotized medially, tergite 7 relatively long, dorsal length ca. 0.9 times epandrial dorsal length; sternite 6 and 7 separated into right and left portions, sternite 6 moderately long, left portion moderately sclerotized, more heavily sclerotized along proximal margin, right portion moderately sclerotized; right portion of sternite 7 relatively long, slightly shorter than right portion of sternite 6, with narrow connection on right to right ventral margin of tergite 7 (this connection posterior to right spiracle 7). Epandrium relatively small, with scattered setulae, 1–2 in posteroventral corner often very long and thick, and long, bare (free of microtrichia and setulae) posteroventral extensions, these fused medially posterior to cerci, posterior margin of sclerotization ending at distinct rough patch proximal to surstyli; surstylus relatively small, separated from epandrium by sclerotized rough patch, ventral face and margins minutely setulose, slightly concave directly opposite of postgonite ventrally, in lateral profile ventral margin rounded, with sharp dorsal bend distally, posterodorsal margin slightly rounded, in posterior profile lateral margin slightly rounded, posteroventral margin concave on distal half, medial margin slightly concave. Cercus elongate, ventral margin flat, dorsal margin ventrally sloping, longest cercal setulae longer than all epandrial setulae but the enlarged posterolateral setulae (when present). Hypandrium with low, slightly angular dorsobasal lobe, posterior bridge ventrally produced, hypandrial arm bending ca. 45˚ at hind margin of epandrium, with 2 ventrolateraly oriented setulae distal to bend and anteriorly oriented process on medial margin distally, not fused to opposite hypandrial arm apically; postgonite with low subapical dorsal process, minutely setulose dorsally. Phallapodeme with large “fan”, margin opposite fan convex basally, concave distally, basiphallus weakly sclerotized basally, elongate, straightening apically; distiphallus base short, ventrally projecting, with wrinkles and coarse pebbled texture dorsally, apex bilobed, lobe bases stacked dorsoventrally, ventral lobe much longer than dorsal lobe, with ventrally produced, moderately sclerotized process with short, broad-based antrorse spinules, otherwise elongate, extending slightly left, heavily sclerotized on right margin, membranous left margin, with broad-based, antrorse spinules on right and fine spinules basally on left, apex bent ventrally and to left, dorsal lobe well sclerotized dorsally, sclerotized area with short, erect, fine spinules, otherwise smooth, membranous, apex acute. Ejaculatory apodeme outside of basiphallus, expanded and with small pores basally.
Female terminalia: Similar to C. flavisetum except as follows. Sternite 5 length 2.6–3.6 times width, sternite 6 length 1.3–1.9 times width. Sternite 8 ca. 2.5 and 3.2 times longer than wide. Spermatheca papillose to papillose/ rugose, papillae broad tipped bumps. Ventral receptacle neck squat, bent at base of donut-shaped apex, duct with fine longitudinal wrinkles.
Comments. One female specimen from Cururipe, Bahia, Brazil, has only one dark seta on its left mid coxa and two dark setae on its right mid coxa. This specimen is identified as C. taeniatum as it has a single dark seta in the posteroventral corner of each katepisternum, its ventral receptacle has a short “neck”, its spermathecae are coarsely papillose, its sternite 8 is 2.5 times longer than wide, and it was collected in the same locality and year (though one month later) as a male C. taeniatum specimen.
Most other female specimens examined fit well into the descriptions given for the three species in the C. taeniatum species complex. Some overlap likely exists between C. taeniatum and C. tumidum sternite 8 proportions and spermathecal texture. Also, the shape of the ventral receptacle varies somewhat within specimens of these two species, and it is plausible some specimens could overlap with the shape found in C. flavisetum . A female specimen from Fazenda Rancho Grande, Rondonia, Brazil, has an elongate sternite 8 (length/width = 3.3), coarsely papillose spermatheca, and a fairly elongate and bent ventral receptacle neck (though not as elongate as in the specimens of C. flavisetum ). This specimen has dark vibrissae, a dark seta at the posteroventral corner of the katepisternum, numerous dark mid coxal setae, and a mix of pale and dark setulae on the anterior surface of the fore femur. It cannot be confidently identified as either C. tumidum or C. taeniatum and, since it is not associated with any male specimens, it has been left unidentified. This problem specimen suggests that the genital characters used to discern females in this complex might have some overlap between species; however this can only be resolved through more extensive sampling.
Curtonotum taeniatum is a junior synonym of Musca gibba Fabricius. The type series of M. gibba Fabricius consists of five syntypes, one of which is a female agreeing with C. taeniatum (this is discussed further below). This specimen is here designated the lectotype of M. gibba Fabricius. The other four M. gibba Fabricius syntypes consist of a male and female of an undescribed species in the C. murinum species complex (we have seen specimens of the same species from Brazil, Suriname, Guyana, and Venezuela) and two C. pantherinum females. The M. gibba Fabricius- C. taeniatum synonymy was first noted by Malloch (1930). He indicated that C. taeniatum agreed withn a female syntype of M. gibba Fabricius housed at the Copenhagen Museum and he therefore treated Curtonotum gibbum as the senior synonym. Malloch commented that the species has a “whitish-dusted central vitta”, a reference almost certainly to the finely pebbled medial frontal vitta unique to the C. taeniatum species complex, so while Malloch does not explicitly state that he examined the type series of C. taeniatum , it certainly appears that he knew what the species looked like. This synonymy was followed by Curran (1934) and Wirth (1975), though Wirth did not indicate which name was the junior and which name was senior. Musca gibba Fabricius is preoccupied by Müller, 1776, and as such C. taeniatum is now the valid name for Musca gibba Fabricius ( Thompson & Pont, 1994) .
While this synonymy has been accepted by multiple authors, none has scrutinized its legitimacy. There are two factors that have to be considered in assessing this synonymy; i) whether or not C. taeniatum and M. gibba Fabricus are actually the same species; and ii) Hendel’s 1933 rebuttal where he rejected the synonymy.
Given that the M. gibba Fabricius lectotype (here designated) is an undissected female, it could be the same species described as C. tumidum by Enderlein. Curtonotum flavisetum can be discounted as a synonym because the lectotype has a black posteroventral katepisternal seta, mostly black medial mid coxa setae, and “reddish brown black” setulae on the anterior face of the fore femur (Kirk-Spriggs, pers. comm.). Dissecting the Musca gibba Fabricius lectotype may allow for more definitive identification, however, given the variability of female genitalic structures, even this might not be conclusive. The origin of the M. gibba Fabricius lectotype is simply given as South America, so neither distributional nor morphological evidence suggests M. gibba Fabricius is more likely synonymous with C. tumidum than C. taeniatum , and there is no reason to consider the synonymy incorrect.
Hendel (1933) refuted the synonymy of C. taeniatum with M. gibba Fabricius , asserting that the first detailed redescription of M. gibba Fabricius was provided by Wiedemann (1830), that Wiedemann’s redescription should be used to identify M. gibba Fabricius , and that Wiedemann’s redescription does not agree with C. taeniatum . Hendel acknowledged that the M. gibba Fabricius type series is a mixture of species, and he contended that Wiedemann (1830) alluded to this by mentioning a second variety of M. gibba Fabricius at the end of his M. gibba Fabricius redescription. Hendel stated it is this second variety that Malloch is synonymizing with C. taeniatum , and that even if C. taeniatum is conspecific with a female specimen in the Fabricius type series, C. taeniatum should still be accepted as a new species as it was only a variety mentioned by Wiedemann, not the species that Wiedemann describes in detail. Based on his own examination of material at Vienna, Hendel stated that the specimens upon which Wiedemann based his description are a mixture of three species: C. taeniatum plus two additional species, one of which is from Brazil. Hendel maintained that his interpretation of M. gibba Fabricius is based on the detailed description that Wiedemann provided and that he used the specimens from Brazil as examples.
While there is logical reasoning in Hendel’s rebuttal, it was certainly not evident in his original treatment (Hendel, 1913) that he was describing a second species from the M. gibba Fabricius type series. Also not clear is whether or not Hendel ever actually saw the Fabricius type series, and whether or not the specimens from Brazil that Wiedemann identified as M. gibba Fabricius and upon which Hendel based his description of M. gibba Fabricius actually agree with any of the specimens in the Fabricius type series.
Malloch’s (1930) decision to synonymize C. taeniatum with M. gibba Fabricius may have been disagreeable to Hendel, but it was certainly not done in error. Malloch made it clear that there is a female type specimen that agrees with C. taeniatum , and that there is a male specimen in the Fabricius type series that does not agree with C. taeniatum (why Malloch assumed that there were only specimens in the M. gibba Fabricius type series unclear). Malloch contended that it is C. taeniatum that Wiedemann (1830) described, pointing out that Wiedemann described the frons as having a pair of blackish spots on the anterior margin. Specimens of C. taeniatum examined in this work do not normally have dark markings on the frons, and in those that do it is assumed to be staining. Other parts of Wiedemann’s description, such as the blackish brown scutum and yellowish femora and tibiae with brown apices, do not match C. taeniatum , and strongly suggest that Hendel was correct in arguing that Wiedemann’s description did not refer to C. taeniatum . However, just what Wiedemann was describing is a moot point as Malloch synonymized C. taeniatum with a female in Fabricius’ type series, a synonymy that, as mentioned above, is likely correct. Examination of the specimens that Hendel and Wiedemann refer to might help further clarify the situation but these specimens (in NHMW) were not available to us.
Given the information currently available and the fact that Curran (1934), Wirth (1975) and Thompson & Pont (1994) followed Malloch’s 1930 synonymy of C. taeniatum and M. gibba Fabricius , it seems most appropriate to continue to treat C. taeniatum as a junior synonym of, and a valid name for, M. gibba Fabricius.
Giglio-Tos (1895), and subsequently Aldrich (1905), reports C. taeniatum (under the names Diplocentra gibba and Cyrtonotum gibbum , respectively) from Tuxpan, Mexico. This is almost certainly based on a misidentification since no Curtonotum species are known to range from Mexico to South America.
Macquart (1843) redescribes C. taeniatum (under the name C. gibbum ) in his description of Curtonotum . Included in the description is mention of a scutellum with six setae, a note that females appear more darkly coloured than males, and two drawings, one of a dorsal habitus, and one of a wing with no infuscation. The variability in appearance is almost certainly because multiple species are involved, not sexual dimorphism. A scutellum with six setae is either a species from the C. murinum species complex, C. apicale , or C. trypetipenne , though the latter is unlikely as the strong wing patterning would be mentioned. The habitus drawing shows six setae on the scutellum but shows a wing similar to that of species in the C. taeniatum species complex species. The wing drawing with no infuscation was likely made from a species in the C. murinum species complex, though not the species included in the Musca gibba Fabricius type series.
Enderlein (1917) provided a wing description for C. taeniatum specimens (under the name Diplocentra gibba ) from Suriname and four Brazilian localities (Espíritu-Santo, Santa Catarina, Minas Gerais and Rio Grande do Sul). The description mentions infuscation on the distal three quarters of cell r 1 and the proximal half of r 2+3. Given the description and the known ranges of possible species Enderlein’s Suriname material is most likely either C. pantherinum or an undescribed species in the C. murinum species complex (the species conspecific with the two M. gibba Fabricius type specimens that belong to the same complex). Enderlein’s Brazilian material is likely C. pantherinum , C. magnum , C. desperatum , C. gracile , C. adusticrus , C. punctithorax , C. papillatum , C. hunkingi , or C. atlanticum .
Type material. The Fabricius type series consists of five specimens. As summarized above, one female agrees with C. taeniatum , a male and female agree with an undescribed species in the C. murinum species complex, and two females agree with C. pantherinum . The female agreeing with C. taeniatum is here designated as the lectotype to avoid any further confusion regarding the definition of M. gibba Fabricius. The C. taeniatum type series includes one male and two females from the same locality in Peru. Hendel (1913) only mentions the two female specimens (neither are designated the holotype in his publication), however it is the male specimen that bears a typus label (the two female specimens do not bear any typus or cotypus labels [Kirk-Spriggs, pers. comm.]). Using this combination of published and unpublished data, these three specimens are here considered the type series (following Article 72.4.1.1, International Code of Zoological Nomenclature , Ride et al., 1999). Since it is clear Hendel intended the male specimen to be the type specimen it is here designated the lectotype. The female specimens are thus designated paralectotypes. A lectotype of C. taeniatum has been designated to avoid any confusion surrounding the definition of C. taeniatum should the original type series prove to be made of more than one species .
Aldrich, J. M. (1905) A catalogue of North American Diptera. Smithsonian Institution, Washington, 680 pp.
Curran, C. H. (1934) The families and genera of North American Diptera. Ballou Press. New York, 512 pp.
Enderlein. G. (1917) Dipterologische Studien. XVI. Dipterologische Notizen. Zoologischer Anzeiger, 49, 57 - 71.
Fabricius, J. C. (1805) Systema antliatorum secundum ordines, genera, species adiectis synonymis, locis, observationibus, descriptionibus. Carolum Reichard, Brunsvigae [= Brunswick], xiv + 372 pp.
Giglio-Tos, E. (1895) Ditteri del Messico. Parte IV. Muscidae Calypteratae-Muscidae Acalypteratae. C. Clausen, Torino, 73 pp.
Hendel, F. (1933) Ueber einige Typen Wiedemann's und Schiner's von acalyptraten Musciden aus Sudamerika, nebst einigen verwandten Arten (Dipt.). Revista de entomologia. Rio de Janeiro, 3 (1), 58 - 83.
Macquart, J. (1843) Dipteres exotiques nouveaux ou peu connus. Memoires de la Societe royale des sciences, de l'agriculture et des arts de Lille 1842, 162 - 460.
Malloch. J. R. (1930) Exotic Muscaridae (Diptera). - XXX. Annals and Magazine of Natural History, 10 (6), 321 - 334.
Muller, O. F. (1776) Zoologiae Danicae prodromus, seu animalium Daniae et Norvegiae indigenarum characteres, nomina, et synonyma imprimis popularium. Typis Hallageriis, Havniae [= Hallager], XXXII, 274 pp.
Ride, W. D. L., Cogger, H. G., Dupuis, C., Kraus, O., Minelli, A., Thompson, F. C. & Tubbs, P. K. (Eds.). (1999) International Code of Zoological Nomenclature. 4 th ed. International Trust for Zoological Nomenclature, London, 306 pp.
Thompson, F. C. & Pont, A. C. (1994) Systematic database of Musca names (Diptera). A catalog of names associated with the genus-group name Musca Linnaeus, with information on their classification, distribution, and documentation. Theses Zoologicae, 20, 1 - 221.
Wiedemann, C. R. W. (1830) Aussereuropaische zweiflugelige Insekten 2: Schulzichen Buchhandlung, Hamm, 684 pp.
Wirth, W. W. (1975) 78. Family Curtonotidae. In: Papavero, N. (Ed.), A catalogue of the Diptera of the Americas south of the United States, Departmento de Zoologia, Sectretaria da Agricultura, Sao Paulo, pp 78.1 - 78.5.
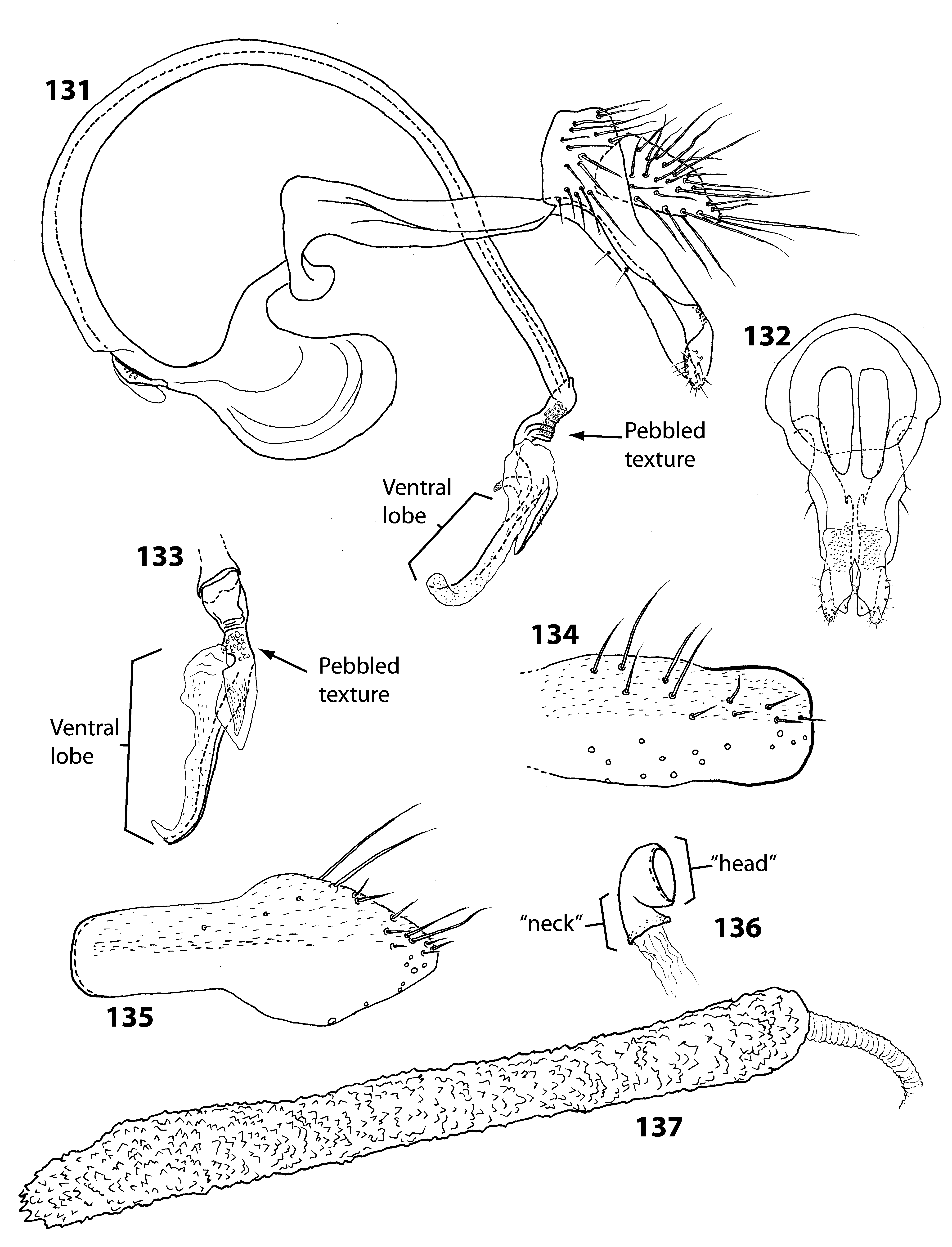
FIGURES 131–137. Curtonotum taeniatum Hendel. 131. Male terminalia, left lateral. 132. Male terminalia, posterior. 133. Distiphallus, dorsal. 134. Female sternite 8, ventral. 135. Female sternite 10, ventral. 136. Ventral receptacle. 137. Spermatheca.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Curtonotum taeniatum Hendel
| Klymko, John & Marshall, Stephen A. 2011 |
Curtonotum gibbum (Fabricius), 1805
| Curran, C. H. 1934: 440 |
| Malloch 1930: 325 |
| Aldrich, J. M. 1905: 641 |
| Macquart, J. 1843: 350 |
Musca gibba Fabricius, 1805: 297
| Fabricius, J. C. 1805: 297 |