
Lygodium hians
|
publication ID |
https://doi.org/ 10.5252/a2014n1a3 |
|
publication LSID |
lsid:zoobank.org:pub:2FDFA93F-3E0F-4669-85CD-349F1BDF1813 |
|
persistent identifier |
https://treatment.plazi.org/id/8E2BF063-C515-FFC9-AA77-FC6F5AD97DA0 |
|
treatment provided by |
Carolina |
|
scientific name |
Lygodium hians |
| status |
|
DESCRIPTION OF LYGODIUM HIANS View in CoL
HABIT AND HABITAT
In the site (discovered by CNP November 1984 and revisited by Dr. Adrian Newton in 2006), Plateau de Dogny in the central mountain backbone of New Caledonia (c. 21° 37’S, 165°50’E, at c. 710 m elevation), the Lygodium colony is present in midmontane rainforest off the ultramafic rocks, on a schistose rock site. The site is in a relatively remote forest-floor habitat ( Fig. 1 View FIG ), with a deep, moist, fine silty forest loam, on a south to south-easterly (hence shaded) aspect, beneath a continuous canopy of tall mid-montane rainforest, and near-to and beneath the canopy of a tree of Austrotaxus spicata Compton. Here , the fronds of this fern were numerous (more than 100), together forming a relatively contiguous frond-colony structure, c. 4-5 m in overall width, of exclusively ground-clothing fronds, across a single patch of forest floor. This montane Lygodium was found on this mountain only in this single location, and no comparable colony was found on several other mountains climbed. At this site, the life-form of this colony, across the period for which it has been known, has been exclusively that of a colony-forming network of ground-clothing fronds. The extent of the colony when first found indicates that it has likely developed slowly but steadily for a considerable period before this, probably decades. Subsequently gathered evidence from previously existing herbarium collections enabled identification of this colony to Lygodium hians E.Fournier. Herbarium specimens typically include either entirely fertile ( Fig. 4B View FIG ), or fertile plus vegetative fronds ( Fig. 4A View FIG ) – the latter serving to firmly make the material-connection with fronds of the type of vegetative structure shown by the Plateau de Dogny colony studied here. The total known range of this species shows its presence on several other New Caledonian high mountains [Mt. Humboldt (type locality, Balansa 1564 ‘in sylvis, 1000 m’; Fournier 1873), Mt. Panié, Mt. Mou, Mt. Arembo, Mt. Colnett, Mt. Dzumac, Nekando, massif de Ton, Massif du Tchingou, Koghis, Puébo, Haute Tchamba, all at altitudes 700-1350 m and especially around 950+ m], with the records most often from single collections ( Brownlie 1969 plus subsequent herbarium data). The majority of the 19th century herbarium specimens of L. hians add little further ecological information, typically beyond that of location and altitude, although several bear the annotation: “très rare”. Perhaps significantly, we have found no previous record of the species for Plateau de Dogny. Indeed, all of the herbarium sheets seen (including those in major European herbaria and those in Noumea Herbarium, and thus representing probably virtually the total that exists), all except two are exclusively of the climbing phase of the plant only, and thus do not recognise the existence of the long-lived vegetative life-phase of the species as described here. Thus with respect to the known herbarium material, had others encountered such purely vegetative material in such mid-montane forest interior locations, it is likely that absence of fertile parts and difficultly in taxonomic attribution of such curious vegetative material, would have discouraged collection of such fronds, and/or their subsequent incorporation into herbaria, through entirely unknown taxonomic affinity. This may explain the apparent lack of any previous description of such morphologically unusual vegetative pteridophyte material. Further, a great majority of the few herbarium specimens located (NOU, P, K, BM, L, E) are from higher-altitude sites (see above), around upper forest limits ( Fig. 4B View FIG ). It is thus to such sites that the occurrence of fertile material would appear to be restricted. Despite the limited herbarium annotations, it is clear that all represent the occurrence of L. hians at such higher altitudes, which would probably be in upper tree-line vegetation, and in cooler and probably lower-growing, more open vegetation, subjected to frequent passing cloud and light precipitation. Indeed, the vegetation type here is of a relatively open forest structure of distinctly temperate character, as indicated especially by the absence at this altitude of lianas, most epiphytes, tree-ferns and palms, all abundant in new Caledonian forests at lower (tropical) altitudes ( Fig. 1A View FIG ). Such high-altitude and temperate forest sites are themselves totally uncharacteristic of most other Lygodium species both in New Caledonia and elsewhere. The location of the patch of L. hians under study on Plateau de Dogny is not only within the overall forest floor community, but is also a relatively mesic, deep forest soil habitat. This is on a sloping mountainside site which is also immediately around and exclusively in close vicinity and over the root-circle of an equally unusual conifer, Austrotaxus spicata . The tree of Austrotaxus spicata was itself the only specimen of its species encountered within the vicinity, and several seedlings of Austrotaxus occurred amongst the fronds of this Lygodium colony ( Fig. 1B View FIG ). Perhaps significantly, this genus is monotypic and endemic to New Caledonia, and the only member of its family ( Taxaceae ) in the southern hemisphere (and is unknown anywhere as a fossil; Page 1990, 2003; Farjon 2008).
SPOROPHYTE MORPHOLOGY
AND SOROPHORE POSITION
Including both herbarium data and those from the site studied in the field, the sporophyte of L. hians differs in structure from that of more typical members of the genus Lygodium in four principal ways:
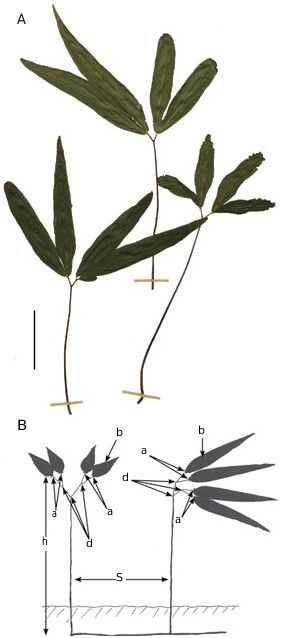
a) Its vegetative fronds are largely or exclusively composed of ones of a definite, rather than indefinite, growth habit, and in the Plateau de Dogny colony of Lygodium hians , the plant shows a predominance, through its life, of a remarkably distinctive frond type ( Figs 1B View FIG ; 2A, B View FIG ). The colony consists of longcreeping slender underground rhizomes, which are c. 2-3 mm in diameter, and occur at a soil depth of c. 10-15 cm and deeper, and which give rise to erect stipes spaced at intervals of c. 5-12 cm or more ( Figs 1B View FIG ; 2B View FIG ). Each stipe, at its top, bears directly a single or usually repeated pair of Y-shaped bifurcations ( Figs 2A View FIG ; 5A View FIG ). These bifurcations, and the blades they bear, terminate the growth. There are no arrested false bifurcations involved, and the form of each frond is, thus, of definite, rather than indefinite growth habit, throughout the vegetative colony ( Figs 1B View FIG ; 2 View FIG ) .
b) These dominant ground-clothing fronds have an entirely dichotomising blade structure ( Figs 1B View FIG ; 2 View FIG ). Each stipe ascends for 8-20 cm, before forming the two closely successive Y-shaped dichotomies.The first branch is regularly of a wide angle of c. 60 degrees spread, the second the narrower at c. 45 degrees spread ( Figs2 View FIG ; 5A View FIG ). Each half of the second branch is sometimes shortly-stalked or, more often, sessile ( Fig. 5A View FIG ). Each of the second dichotomies usually carries only a single blade, which is obtuse at its base, widening rapidly to about 2.2-2.5 cm, thence tapering gradually over a length of up to 9-18 cm (fide Page 23 221, E). The reduced length of some blades ( Fig. 2A View FIG right frond and right blade pair of centre frond) is due to extensive insect feeding damage ( Fig. 5C View FIG ). Because of the narrower second angle, most fronds appear to be structured as two separated sets of closely-paired long-tapering ‘pinna’- like leafy-segments ( Fig. 2 View FIG ). Across the region of the dichotomies, the frond orientates each of these ‘pinnae’ to an ascending-spreading angle of c. 30- 50° from horizontal. Significantly, the very base of each blade carries a well-formed abscission zone, itself a rare feature in Pteridophyta as a whole. The blades have open dichotomous venation with one or two dichotomies between mid-vein and margin ( Fig. 5B View FIG ). Blade margins are finely denticulate, tooth positions not related to vein endings ( Fig. 5B View FIG ). The overall appearance of the vegetative fronds is so distinctive and unusual that close examination was needed on first encounter to initially establish that this was indeed a fern. Superficially, the fronds have an overall similarity to a dwarf bamboo (such as Sasa ). Fronds with such dichotomising blades thus not only predominate the life-cycle, but also can persist relatively indefinitely through time in this form building substantial colonies as on Plateau de Dogny.
c) These ground-clothing fronds are of a quite dark-blue-green colouration, and have a toughleathery, fairly thick and semi-rigid texture, totally unlike the pale yellow-green colour and thin blade texture more characteristic of the vegetative parts more widely known on climbing fronds of other species of Lygodium .
d) Herbarium data indicate that climbing fronds can be eventually produced, at least by some higheraltitude colonies in New Caledonia. Indeed, two Paris herbarium specimens ( Fig. 4 View FIG ) from Mt. Arembo & Mt. Panié) show both a sterile and fertile frond on the same sheet, the latter specimen with a note (translated): “fern climbing through the bushes or ground-covering, very common but almost always sterile”. (We think the reference to “common” may refer to a local frequency of fronds, as members of a single large clone, similar to the one on Plateau de Dogny). These demonstrate that fertile fronds when formed can climb to 1-4 m high, but such a climbing frond-state may be produced relatively briefly (both in stature and likely duration). Each fertile blade is usually appreciably shorter than the vegetative ones in the ground-clothing fronds studied here, usually 5.0-7.5(-11.5) cm in overall length. Sporangia are produced within specialised sorophores, themselves borne from peg-like protrusions, with the sorophores each placed singly or in pairs at the end of a vein. Laminar protrusions bearing the sorophore clusters are very occasionally few, imperfectly-formed and lateral, but on most specimens are abundant, regularly formed and grouped into continuous close-spaced terminal and immediately sub-terminal positions, thus forming regularly regimented arrays around the apex of each fertile blade ( Fig. 4 View FIG ,including Lectotype P00523232, but demonstrated especially clearly and extensively also on Noumea herbarium sheets Buisson 1425 and Veillon 5660). Thus the distribution of sorophores around the blades differs in L. hians from those of other species of Lygodium , in being mostly terminal on each blade (rather than arrayed in mainly lateral positions along both longer margins as in most other species of Lygodium ).
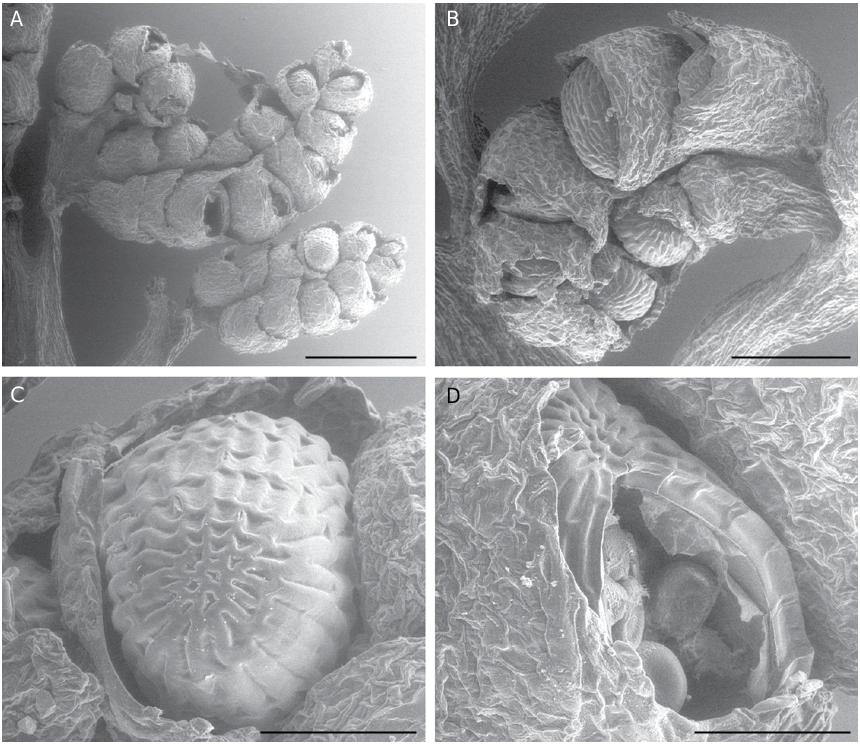
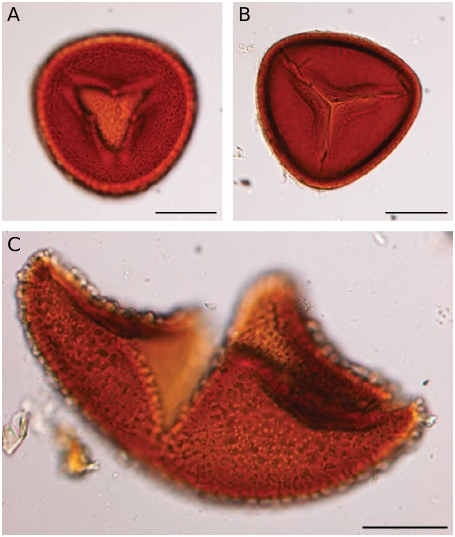
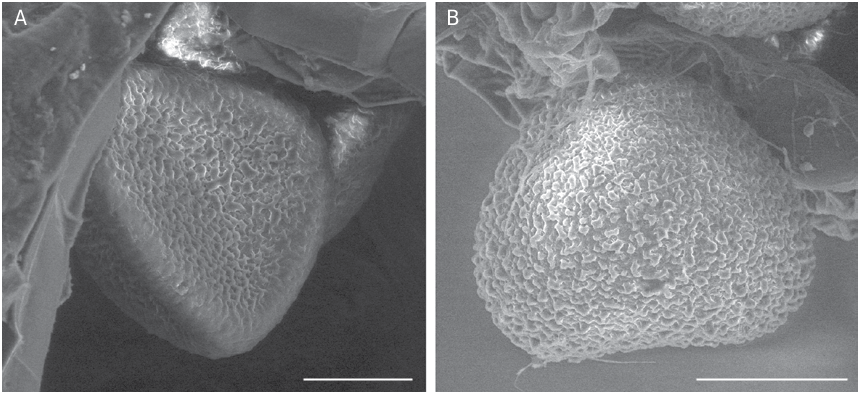
SOROPHORE AND SPORANGIA MORPHOLOGY Sorophores and sporangia of L hians are typical for all Lygodium , characterized by having the sporangia borne in so-called sorophores, in two rows at the end of veins; each row bearing 5-6 sporangia ( Fig. 6A, B View FIG ). Sporangia are borne on marginal lobes covered by a laminar outgrowth (indusium) ( Fig. 6A, B View FIG ). Sporangia in L. hians are ca. 600 µm long ( Fig. 6A, B View FIG ), with a clear apical annulus ( Fig. 6C View FIG ). Longitudinal dehiscence of the sporangia reveals a number of irregularly reticulate to verrucate spores ( Fig. 6D View FIG ). Spores of Lygodium are in general 50-130 µm in diameter, always trilete, often with a margo along the laesurae and have a thick perispore that is usually scabrate, granulate or verrucate; sculptural elements sometimes fused to form a reticulum; the exospore is slightly thinner and more finely sculptured (van Konijnenburg-van Cittert 1991, 1992). The spores of L. hians are no exception, except that they are the largest of all known Lygodium spores: with the perispore they measure between 120 and 150 µm ( Fig. 7A View FIG ); when the perispore is lost ( Fig. 7B View FIG ) they are between 100 and 140 µm. The laesurae are c. 2/3rd of the spore radius, and are accompanied by a clear margo ( Fig. 7B View FIG ). The exospore (up to 5 µm thick) is granulate, granules often forming an irregular, fine reticulum ( Fig. 7A, B View FIG ). Perispore thick (up to 8 µm), coarsely and irregularly verrucate to even rugulate ( Fig. 7C View FIG ); verrucae often fused, forming a coarse, very irregular reticulum ( Fig. 8A, B View FIG ). The spore morphology is character- istic of the Lygodium subgenus Odontopteris (van Konijnenburg-van Cittert 1991, 1992). However, its large spore size might be a derived feature, as a spore size between 50-70 µm is considered as be- ing original in Schizaeales and much larger spore sizes (e.g., in Lygodium and some Schizaea species ) may be derived (see e.g., Van Konijnenburg-van Cittert 1991, 1999).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |