
Solaenodolichopus Verhoeff, 1924
|
publication ID |
https://doi.org/ 10.5852/ejt.2014.83 |
|
publication LSID |
lsid:zoobank.org:pub:049F326B-9460-4038-BB21-9DA36F79812F |
|
DOI |
https://doi.org/10.5281/zenodo.3851834 |
|
persistent identifier |
https://treatment.plazi.org/id/8B242A14-8B7A-4F31-8944-BBBCFAFF978C |
|
treatment provided by |
Tatiana |
|
scientific name |
Solaenodolichopus Verhoeff, 1924 |
| status |
|
Solaenodolichopus Verhoeff, 1924 View in CoL
Solänodolichopus Verhoeff, 1924: 10 (in key as subgenus of Antichiropus View in CoL ), 11 (in key as subgenus of Antichiropus View in CoL ), 19 (first description).
Paraulacoporus Verhoeff, 1928: 88 View in CoL , 90 (in key), 91 (in key), 93. [Type species Paraulacoporus sulcatus Verhoeff, 1928 , by monotypy.]
Parwalesoma Verhoeff, 1937: 139 View in CoL (first description). [New synonymy. Type species Parwalesoma castaneum Verhoeff, 1937 View in CoL , by monotypy.]
Solaenodolichopus View in CoL – Attems 1926: 143 (in key as subgenus of Antichiropus View in CoL ); 1937: 260 (synonymised with Aulacoporus View in CoL ). — Verhoeff 1932: 1574 (as subgenus of Antichiropus View in CoL ), 1595 (in key), 1597 (in key). — Jeekel 1965: 8; 1968: 29 (as synonym of Aulacoporus View in CoL ); 1971: 233 (type species designated); 2000: 37 (in key), 40–42; 2003: 42. — Hoffman 1980: 166 (as synonym of Aulacoporus View in CoL ). — Nguyen & Sierwald 2013: 1159.
Solänodolichopus – Verhoeff 1928: 88 (raised to genus), 90 (in key), 91 (in key), 94. — Brolemann 1931: 299. — Verhoeff 1937: 139. — Verhoeff 1941: 8 (in key).
Paraulacoporus View in CoL – Attems 1929: 258. — Brolemann 1931: 295, 298. — Verhoeff 1932: 1595 (in key), 1598 (in key). — Attems 1937: 260 (synonymised with Aulacoporus View in CoL ). — Verhoeff 1941: 8 (in key). — Jeekel 1965: 8. — Jeekel 1968: 29 (as synonym of Aulacoporus View in CoL ). — Jeekel 1971: 230 (in genus catalogue). — Hoffman 1980: 166 (as synonym of Aulacoporus View in CoL ). — Jeekel 2000: 40, 42 (synonymised with Solaenodolichopus View in CoL ). — Nguyen & Sierwald 2013: 1159 (as synonym of Solaenodolichopus View in CoL ).
Parwalesoma View in CoL – Attems 1940: 549 (Verhoeff’s description reworded). — Verhoeff 1941: 8 (in key). — Jeekel 1965: 8. — Jeekel 1968: 29, 126. — Jeekel 1971: 231. — Hoffman 1980: 166. — Jeekel 2000: 37 (in key), 40-42. — Jeekel 2003: 42. — Nguyen & Sierwald 2013: 1157.
Type species
Antichiropus (Solaenodolichopus) teres Verhoeff, 1924 View in CoL , by subsequent designation.
Other assigned species
S. pruvoti ( Brolemann, 1931) , S. rubriventris Verhoeff, 1928 , S. sulcatus ( Verhoeff, 1928) , S. vittatus ( Verhoeff, 1924) and S. walesius Verhoeff, 1928 .
History of Solaenodolichopus
The sometimes confusing development of the Solaenodolichopus concept over the past 90 years has involved five genus names and four diplopodologists.
Besides Solaenodolichopus , some of the species redescribed in this paper have also been assigned to Antichiropus Attems, 1911 , Aulacoporus Verhoeff, 1924 , Paraulacoporus Verhoeff, 1928 and Parwalesoma Verhoeff, 1937 .
The four diplopodologists were the Austrian Carl Attems (1868–1952, in full, Carl Aug. Graf [Count] Attems-Petzenstein), the German Karl Wilhelm Verhoeff (1867-1944), the Frenchman Henry Wilfred Brölemann (1860-1933) and the Dutchman Casimir Albrecht Willem Jeekel (1922-2010). I use the spelling ‘Brolemann’ below rather than ‘Brölemann’, following that author’s practice in the 1931 publication cited in this paper. Brolemann dropped the umlaut in his name from 1920 onwards (J.-J. Geoffroy, in litt., 30 Apr. 2009).
All four specialists viewed the polydesmidan gonopod as a modified walking leg on which it was possible to recognise podomere homologs. Their differing opinions on homology are mentioned only briefly below in the context of a chronology of Solaenodolichopus . My purpose here is to try to explain the reasoning behind particular generic assignments in the past, not to thoroughly review an enduring and still-unresolved controversy.
Solaenodolichopus was first proposed by Verhoeff (1924) as a subgenus of Antichiropus Attems, 1911 . Attems (1911) had described seven new species from Western Australia with gonopod telopodites divided into three main parts: a small, densely setose, basal portion (Femur); a long, cylindrical, slender, central portion (Tibia) tipped with short prongs (Zacken); and a long, thin, coiled, apical portion (Kanalast) carrying the prostatic groove and its terminal opening. Attems assumed that the tarsus of the modified leg had fused with the tibia, and that the distal portion of the tibia was its tarsal section (Tarsalabschnitt). In his view, the first joint of this tarsal section consisted of the Kanalast and a prong-like process ‘a’ at the end of the tibia and close to the prostatic groove. Attems suggested that another tibial process, ‘c’, was a remnant of a second tarsal joint. (For a discussion of the Antichiropus gonopod and its components, see Car et al. 2013.)
Verhoeff (1924) described the telopodites of the new central Queensland species teres and the southern Queensland vittatus (the latter divided into two subspecies) as likewise having a small, densely setose, basal portion (Praefemur); a long, cylindrical, slender, central portion (Femur) tipped with short accessory branches (Nebenäste); and a long, terminal process carrying the prostatic groove and its opening. Verhoeff regarded the latter as an evolutionarily novel structure (Neubildung) he called a Solänomerit, which he had recognised in many other Polydesmida and which had no homologue on a walking leg; in what follows I will use the modern term ‘solenomere’. The two Queensland species were placed in Antichiropus (Solaenodolichopus) not because this arrangement was definitive (wrote Verhoeff), but because the two species seemed more closely related to those in Antichiropus than to any other species he had previously examined. The gonopod telopodites were fundamentally similar and the first male legs (bearing a short, rounded, ventral, femoral process) were almost identical.
According to Verhoeff (1924), the polydesmidan solenomere was primitively protected by a long tibiotarsus, but there were four secondary developments away from this pattern. In one development (Verhoeff’s case 1), the tibiotarsus was reduced so much that the solenomere became the free end of the gonopod, as in Antichiropus . The tibiotarsus had either disappeared completely in Antichiropus , or had been reduced to one of the small accessory branches at the end of the femur (Attems’ Zacken). In another development (case 4), the tibiotarsus and solenomere fused into a single process. The two developments could overlap ( Verhoeff 1924: 6, “Die Fälle N. 1 und 4 gehen übrigens in einander über”). In the new Queensland species, Verhoeff believed he could detect a fusion in which the solenomere had dominated, so that the tibiotarsus was reduced to a small subterminal tooth on the teres solenomere and a larger but likewise distal process on the solenomere in vittatus .
In the same 1924 paper, Verhoeff proposed Aulacoporus as a new genus for three northern Queensland species. Aulacoporus was distinguished from Antichiropus (Solaenodolichopus) in two ways. First, the prostatic groove opened at the extreme tip of the solenomere in Aulacoporus , whereas in Antichiropus (Solaenodolichopus) the opening was carried on a near-apical process of the solenomere. The solenomere and tibiotarsus had fused in Aulacoporus , but the tibiotarsus had completely disappeared in one Aulacoporus species and in the other two was reduced to a distal bulge on the solenomere.
To explain Verhoeff’s second diagnostic difference between Aulacoporus and Antichiropus (Solaenodolichopus) , I need to digress briefly.
Polydesmidan species have been classified, in part, on the structure of the paranota, which are lateral extensions of the body wall. In many families the paranota are wide, dorsoventrally flattened and more or less horizontal. In some families there are species with no detectable paranota, at least on diplosegments, i.e. the body rings are simple circles in cross-section. Between these two extremes, paranota are often said to be ‘reduced’. Verhoeff (1924) attempted to refine the description of paranota for classification purposes. Instead of Kiel (keel, carina) for paranota in general, Verhoeff proposed Seitenwülste (lateral bulges) for all lateral swellings and extensions. In species with wide, dorsoventrally flattened, wing-like Seitenwülste, the term Seitenflügel (lateral wings) would be appropriate. Each Seitenwulst was often accompanied medially by a narrow, dorsal, longitudinal furrow which Verhoeff called a Seitenfurche (lateral groove). Species with no Seitenwülste at all might or might not have Seitenfurchen.
Attems (1937) used these terms slightly differently. He observed that well-developed paranota, i.e., Seitenflügel, often had swollen lateral margins. In some species the paranota had narrowed so much that only these marginal thickenings were still present, in which case one could speak of Seitenwülste, and in species without Seitenwülste there might still remain the medial, demarcating grooves, the Seitenfurchen. Both Attems and Verhoeff understood these variations as an evolutionary series in which primitively wide paranota were progressively reduced to bulges and, before disappearing entirely, to short, fine, lateral grooves on the metazonites.
The Aulacoporus and Antichiropus (Solaenodolichopus) species described by Verhoeff had reduced but detectable paranota on the haplosegments, i.e. rings 2-4. Neither group had Seitenwülste on the diplosegments, but there were distinct Seitenfurchen in Aulacoporus and none at all in Antichiropus (Solaenodolichopus) .
Attems (1926) accepted both Aulacoporus and Solaenodolichopus (as a subgenus of Antichiropus ), but reiterated his view that beyond the coxa on the gonopod were only a femur, tibia and tarsus. Verhoeff’s Solänomerit was simply a tibial extension (Tibialfortsatz). In a key to paradoxosomatid genera (‘Strongylosomidae’), Attems (1926) separated Aulacoporus (and other genera) having a simple telopodite without side branches or a tarsal section, from Antichiropus (and other genera) having a multitipped telopodite. The key listed Antichiropus as having 1-3 short processes, probably tarsal remnants, near the base of the tibial extension (i.e., the solenomere).
Verhoeff (1928) raised Solaenodolichopus to a genus and added two new species from NSW, rubriventris and walesius . He also proposed Paraulacoporus for another new NSW species, sulcatus . In Paraulacoporus as in Aulacoporus there were short Seitenfurchen on most diplosegments, although the grooves were indistinct on rings 15-19. Unlike Aulacoporus but like Solaenodolichopus , Paraulacoporus was characterised by a prostatic groove opening proximal to the solenomere tip. Unlike the situation in Solaenodolichopus , the two accessory branches at the end of the femur in Paraulacoporus were of equal size, and the process lateral to the prostatic groove opening on the solenomere was forked. Verhoeff described this forked process as a ‘tarsal branch’.
Brolemann (1931) assigned a new New Caledonian species, pruvoti , to Paraulacoporus , although he observed no lateral groove (sillon) past ring 5. Combining the ideas of Attems and Verhoeff, Brolemann described the pruvoti telopodite as consisting of a short, densely setose, basal femoroid (fémoroïde), a long, slender tibia and a terminal seminal branch (rameau séminale). He regarded the short, posteromedial process at the end of the tibia as a reduced tarsus separate from the seminal branch, and rejected Verhoeff’s opinion that the seminal branch in P. sulcatus was a fusion of solenomere and tibiotarsus. Brolemann also objected to the placement of Solaenodolichopus within Antichiropus by Verhoeff and Attems. He suggested instead that Paraulacoporus and Solaenodolichopus could be grouped together because both had an erect, expanded, more or less divided seminal branch, not a long, coiled strap as in Antichiropus .
Verhoeff (1932) offered simple new keys to the known Australian paradoxosomatids (‘Strongylosomidae’) based either on trunk details or gonopod structure. In the latter key, Aulacoporus , Paraulacoporus , Solaenodolichopus and Walesoma Verhoeff, 1928 were grouped as genera in which a long solenomere led from the end of the femur, which also might have 1-2 accessory branches at its apex. Aulacoporus and Walesoma were paired as genera in which the opening of the prostatic groove was at the extreme end of the telopodite, with no accessory process either proximal or distal to the opening. Paraulacoporus and Solaenodolichopus were paired as genera in which the opening of the prostatic groove was proximal to the extreme end of the telopodite, with accessory processes both proximal and distal to the opening.
Attems (1937) revised his nomenclature of sections of the gonopod telopodite, renaming femur, tibia and tarsus as prefemur, femur and tibiotarsus. The process carrying the prostatic groove (Samenrinne) and its opening was the Rinnenast (groove-branch) and was a process of the femur. More importantly, in 1937 Attems synonymised both Paraulacoporus and Solaenodolichopus under Aulacoporus . He described the gonopod and paranota in the expanded genus as follows ( Attems 1937: 260-261, my translation): “Gonopod femur long to very long and slender, at the end with one or no side-branch. Rinnenast very large and broad, with one, sometimes two side-teeth. The tibiotarsus is reduced to a small spine. Seitenwülste weakly developed, low, round, sometimes only present on anterior segments. Longitudinal grooves either present on most segments or lacking from segments 5 or 6 onwards”. In his key to Aulacoporus species, Attems separated the former Aulacoporus species from the former Paraulacoporus and Solaenodolichopus species simply on the presence or absence of Seitenfurchen on most rings.
Verhoeff (1937) described the new species castaneum from NSW and assigned it to a new genus, Parwalesoma . He noted that Parwalesoma was close to both Aulacoporus and Solaenodolichopus , but differed from the former genus in lacking Seitenfurchen on most rings. Parwalesoma and Solaenodolichopus differed in a somewhat sharper demarcation between femur and solenomere, and in the solenomere tip in P. castaneum being spoon-shaped. Parwalesoma castaneum Verhoeff, 1937 was in fact Solaenodolichopus walesius Verhoeff, 1928 redescribed from new material (see below under S. walesius ), but it was another 60+ years before Verhoeff’s confusion was detected by Jeekel (2000; see below).
In his last contribution to the Solaenodolichopus story, Verhoeff (1941) unwittingly redescribed Brolemann’s S. pruvoti (from New Caledonia) as Solaenodolichopus annulatus (from Brisbane). He also offered a key to genera in which he separated the taxa lumped by Attems (1937). The relevant portion of the key is as follows ( Verhoeff 1941: 8, my translation with Verhoeff’s terminology updated):
c) Solenomere divided at the end into two branches, the medial one extending as three protuberances, of which the middle contains the opening of the prostatic groove, the proximal one a bend in the prostatic groove. Trunk without Seitenwülste but with Seitenfurchen …… Paraulacoporus Verh.
d) Prostatic groove without bend before its end …………………………………………………e, f
e) Solenomere before its end with a spoon-shaped concavity. Trunk with neither Seitenwülste nor
Seitenfurchen …………………………………………………………………… Parwalesoma Verh. f) Solenomere without this concavity ……………………………………………………………g, h
g) End of the solenomere drawn out into two closely standing tips, of which the more proximal contains the opening of the prostatic groove. Trunk with neither Seitenwülste nor Seitenfurchen …………………………………………………………………………… Solaenodolichopus Verh.
h) Opening of the prostatic groove in a simple, apical tip……………………………………………i, j
i) Solenomere significantly shorter than femur, its base enlarged, its end hooked. Trunk without Seitenwülste but with Seitenfurchen ………………………………………… Aulacoporus Verh.
j) Solenomere and femur about equally long, the solenomere base not swollen. Trunk with weak, posteriorly rounded Seitenwülste and Seitenfurchen ……………………………… Walesoma Verh.
Reviewing the Australian Antichiropodini, Jeekel (1968) accepted Attems’ (1937) synonymies and Verhoeff’s Parwalesoma . Thirty-two years later, he confessed that he regarded Parwalesoma as another Aulacoporus synonym, but had not published this synonymy because the Parwalesoma type species, P. castaneum Verhoeff, 1937 , would have become a junior homonym of the Aulacoporus type species, A. castaneus Verhoeff, 1924 . “As the broad concept of Aulacoporus was intended to be a temporary
solution anyway, the introduction of a new name for the type-species of Parwalesoma seemed just too radical” ( Jeekel 2000: 40).
Jeekel (2000) then reinstated Solaenodolichopus , leaving within it S. teres , S. vittatus and S. vittatus dorsalis and transferring to it the Paraulacoporus species P. sulcatus and P. pruvoti . He also recognised S. annulatus as a synonym of pruvoti . Solaenodolichopus now included species with and without Seitenfurchen, and with considerable variation in the size and position of the processes at the end of the telopodite femur. “The species of this genus are characterised by the solenomerite of the gonopods having a pre-apical latero-distal process paralleling the apex proper. This process distinguishes the genus from the others” ( Jeekel 2000: 41).
One of those other genera was Aulacoporus and its three original species, which Jeekel (2000) regarded as distinct not because the solenomere was simple with the prostatic groove opening at its tip, but because the gonopod apex curved strongly cephalad and bore a large lobe laterally – a lobe which Jeekel presumed was the tibiotarsus.
The other genus Jeekel (2000) distinguished from Solaenodolichopus was Parwalesoma . He recognised the type species P. castaneum as a junior subjective synonym of S. walesius , thus establishing P. walesium ( Verhoeff, 1928) . He also transferred S. rubriventris to Parwalesoma , noting that “In this genus the gonopods are lacking a pre-apical process on the solenomerite, but otherwise are quite similar to those of Solaenodolichopus ” ( Jeekel 2000: 42).
Jeekel’s distinction between Parwalesoma and Solaenodolichopus is questionable in the case of S. walesius (see the taxonomic notes following the S. walesius redescription, below), and for that reason I am making Parwalesoma a junior subjective synonym of Solaenodolichopus . This action broadens the circumscription of Solaenodolichopus and might allow it to be subsumed under Aulacoporus , as favoured by Attems (1937). However, after examining late 20th-century specimens of Aulacoporus castaneus Verhoeff, 1924 , A. yarrabahnus Verhoeff, 1924 and three undescribed Aulacoporus spp., all from a small area in the Queensland Wet Tropics, I agree with Jeekel (2000) that this genus is best kept separate from Solaenodolichopus . All Aulacoporus forms have prominent lateral grooves and slight paranotal swellings on all diplosegments, and in four of the six Aulacoporus forms the solenomere curves anteriorly. None of the Solaenodolichopus I have examined (as diagnosed above) has either prominent lateral grooves on all diplosegments or an anteriorly curving solenomere.
Diagnosis
Australian Paradoxosomatidae without distinct paranota on posterior rings, and with a gonopod telopodite with a small, densely setose prefemur and a more or less straight, slender and cylindrical femorite ending in an unbranched medial process, an unbranched lateral process and a much larger solenomere with a flattened tip on which the prostatic groove usually opens medially.
Description
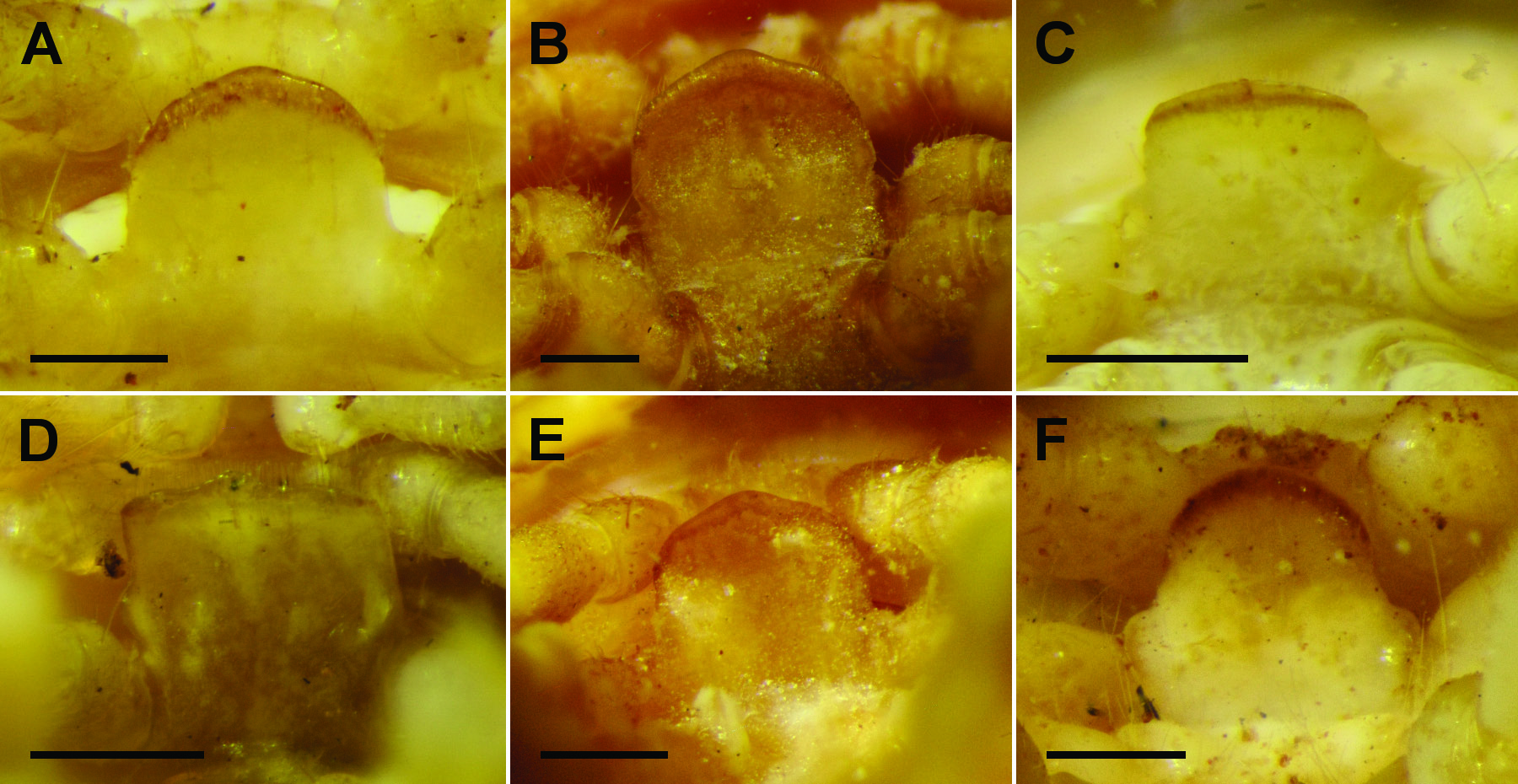
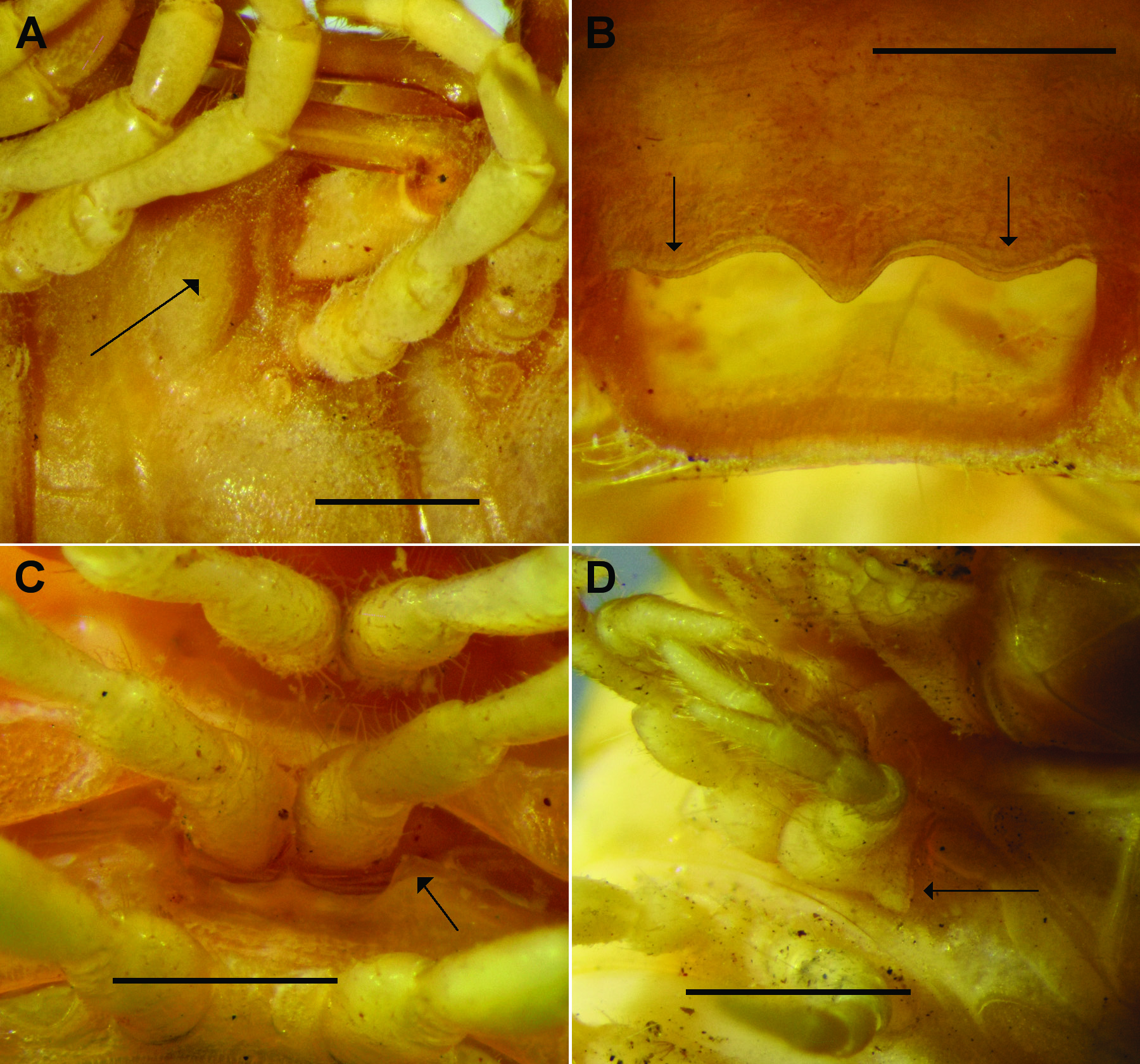
Male with vertex bare, frons and clypeus sparsely setose; vertigial sulcus distinct, ending anteriorly just above level of antennal sockets; post-antennal groove shallow; antennal sockets separated by 1.0-1.1X a socket diameter. Antenna filiform ( Fig. 1A View Fig ), reaching dorsally to rear of ring 2; antennomere with relative lengths typically (2,3,4)> (5,6), the differences small, and maximum distal widths subequal or with 6 very slightly wider. Head and collum about equal in width in dorsal view, about as wide as rings 2–4 or a little narrower; diplosegments a little wider, slightly increasing in width to midbody, width decreasing progressively on last rings. Collum D-shaped in dorsal view; anterior margin straight medially, gently curving laterally; posterior margin straight or very slightly emarginate medially; collum corner broadly rounded ( Fig. 1A View Fig ). Ring 2 paranotum ( Fig. 1A View Fig ) a slightly thickened, overhanging ridge, nearly straight and more or less longitudinal, corners rounded, set at ca. 1/4 ring height or a little less, below level of collum corner. Rings 3 and 4 paranota ( Fig. 1A View Fig ) at about level of collum corner, reduced to shallow furrows curving upwards anteriorly. Pleural keels present on rings 2–4 as slight thickenings with upwardly curving margins. Diplosegments without paranotal thickenings, rings circular in crosssection. Prozonites and metazonites smooth, bare; waist short, shallow, with suture at anterior edge, sculptured indistinctly as variably low, rounded, longitudinal ridges; limbus a short, thin, continuous sheet. Pore formula normal; ozopores small, round, flush with lateral surface of ring, set at mid-height and mid-length on metazonite, in line with transverse furrow (when present). Diplosegment spiracles ( Fig. 1B–C View Fig ) just above and anterior to leg bases, more or less egg-shaped with smaller end ventral, anterior spiracle larger and extended anteroventrally; spiracles with thin, short rim; spiracular filter slightly emergent, forming rounded fold in inverted, tight U-shape in spiracular opening ( Fig. 1B–C View Fig ). Midbody sternites about as long as wide, sparsely setose; cross impressions wide, shallow, transverse impressions a little narrower and deeper than longitudinal. Leg 1 ( Fig. 1D View Fig ) with small, bluntly rounded, ventral, femoral process curving slightly distally, reaching to ca. 1/2 length of femur on ventral side. Midbody legs with relative podomere lengths femur> prefemur> (postfemur, tibia, tarsus) or tarsus very slightly longer than postfemur and tibia; femur 1.7–1.8X as long as tarsus; prefemora a little swollen dorsally. Dense brush (scopula) ( Fig. 1E View Fig ) of straight, finely pointed setae ventrally on tarsus and distal portion of tibia of most legs. Epiproct extending a little past anal valves, tapering in dorsal view with sides more or less straight, ca. 1/4-1/3 maximum width of preanal ring at apex; apex truncate, corners produced posteriorly as small, rounded bumps ( Fig. 1F View Fig ); spinnerets in rectangular array, wider than long. Hypoproct subtrapezoidal to broadly paraboloid. Gonopore small, round, opening ventrally on slight distomedial swelling of leg 2 coxa. Sternal lamella ( Fig. 2 View Fig ) wide, ca. 80–90% of width between leg 4 bases, perpendicular to ring or slightly tilted anteriorly, with sparse long setae on posterior surface and dense, short setae on anterodistal surface. Aperture ca. 1/3 of ring 7 prozonite width, roundedrectangular and tilted slightly posteriorly; prominent, rounded, transverse swelling just anterolateral to aperture on either side ( Fig. 3A View Fig ).
Female very similar to male but with legs proportionally shorter, prefemora not swollen; epigyne ca. 1/6-1/5 ring width, ends slightly raised and cradling leg 2 coxae, centre slightly raised in small, rounded triangle ( Fig. 3C View Fig ).
Species-diagnostic characters
Adults of the six named Solaenodolichopus species conform to the genus description above. Omitted from the genus description are diagnostic characters reported in the descriptions of individual species, below. Besides gonopod structure, these species-diagnostic characters are: colour, maximum midbody width, transverse furrow distinctness, longitudinal furrow details for diplosegments, distribution of scopulae on legs, sternal lamella details, leg spacing on rings 6 and 8, shape of the anterior margin of the gonopod aperture, and leg 2 coxa details in females.
In the species descriptions I use the terms ‘prefemur’, ‘femorite’ and ‘solenomere’ in agreement with the usage of Car et al. (2013: 84) for Antichiropus ; the terms are not intended to be implied statements of homology.
A brief dichotomous key to the six named Solaenodolichopus View in CoL species follows. The key will need major revision as the many undescribed species are named and documented in future papers in this series.
1. Body pale yellow or light brown with narrow dark brown rings at the rear of each metazonite ………………………………………………………………………… S. pruvoti ( Brolemann, 1931) View in CoL
– Body more or less uniformly dark in colour, with or without a lighter, medial dorsal, longitudinal band …………………………………………………………………………………………………2
2. Longitudinal furrows laterally at level of ozopore on diplosegments to about ring 15, medial and lateral femorite processes obvious, spine-like, about equal in length …… S. sulcatus ( Verhoeff, 1928)
– No longitudinal furrows laterally on any diplosegment, medial and lateral femorite processes greatly different in length or both inconspicuously small …………………………………………3
3. Lateral femorite process much larger than medial ………………… S. walesius Verhoeff, 1928 View in CoL
– Medial femorite process much larger than lateral, or both femorite processes inconspicuously small …………………………………………………………………………………………………4
4. Distal 1/3 of femorite bent at ca. 45°, femorite processes inconspicuously small …………………… ……………………………………………………………………………… S. teres ( Verhoeff, 1924) View in CoL
– Femorite straight, medial femorite process large and obvious ………………………………………5
5. Solenomere with several spine-like processes in basal half, no large indentation in distal half ………………………………………………………………………… S. rubriventris Verhoeff, 1928 View in CoL
– Solenomere without spine-like process in basal half, with large, U-shaped indentation in distal half …………………………………………………………………… S. vittatus ( Verhoeff, 1924) View in CoL

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Strongylosomatidea |
|
Family |
|
|
SubFamily |
Australiosomatinae |
|
Tribe |
Antichiropodini |
Solaenodolichopus Verhoeff, 1924
| Mesibov, Robert 2014 |
Parwalesoma
| Nguyen A. D. & Sierwald P. 2013: 1157 |
| Jeekel C. A. W. 2000: 37 |
| Jeekel C. A. W. 1971: 231 |
| Jeekel C. A. W. 1968: 29 |
| Jeekel C. A. W. 1965: 8 |
| Verhoeff K. W. 1941: 8 |
| Attems C. 1940: 549 |
Parwalesoma
| Verhoeff K. W. 1937: 139 |
Paraulacoporus
| Nguyen A. D. & Sierwald P. 2013: 1159 |
| Jeekel C. A. W. 2000: 40 |
| Jeekel C. A. W. 1971: 230 |
| Jeekel C. A. W. 1968: 29 |
| Jeekel C. A. W. 1965: 8 |
| Verhoeff K. W. 1941: 8 |
| Attems C. 1937: 260 |
| Verhoeff K. W. 1932: 1595 |
| Brolemann H. W. 1931: 295 |
| Attems C. 1929: 258 |
Paraulacoporus
| Verhoeff K. W. 1928: 88 |
Solänodolichopus
| Verhoeff K. W. 1941: 8 |
| Verhoeff K. W. 1937: 139 |
| Brolemann H. W. 1931: 299 |
| Verhoeff K. W. 1928: 88 |
Solaenodolichopus
| Nguyen A. D. & Sierwald P. 2013: 1159 |
| Jeekel C. A. W. 1968: 29 |
| Jeekel C. A. W. 1965: 8 |
| Verhoeff K. W. 1932: 1574 |
| Attems C. 1926: 143 |
Solänodolichopus
| Verhoeff K. W. 1924: 10 |