
Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & García-París, 2023
|
publication ID |
https://doi.org/ 10.5852/ejt.2023.860.2053 |
|
publication LSID |
lsid:zoobank.org:pub:EEA89326-4AF7-47A4-8DB6-751DF1BC5D87 |
|
DOI |
https://doi.org/10.5281/zenodo.7689657 |
|
persistent identifier |
https://treatment.plazi.org/id/8A2E87EF-FFD7-FFB2-F574-9F82E111FC83 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & García-París |
| status |
sp. nov. |
Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & García-París sp. nov.
urn:lsid:zoobank.org:act:62F051DC-0763-4A90-A97B-A5F4E79FFFDC
Figs 3–8 View Fig View Fig View Fig View Fig View Fig View Fig
Diagnosis
Dry-mounted specimens light brown, although the head may remain somewhat orange. In living specimens, anterior part of body orange more or less intense (head, first antennomere, pronotum, tegmina and legs), sometimes with darkened areas ( Figs 4–5 View Fig View Fig ). Antennae light brown, with 12 antennomeres; antennomere 1 almost equally long than 2–3 combined. All antennomeres pubescent; a dense, fine and white pubescence covers from the middle of the third antennomere, less frequently from the fourth, to the apical end. Postfrontal sutures do not reach the eyes generally. Pronotum smooth, 1.2 times as long as wide, parallel-sided or slightly wider posteriorly. Tegmina short, truncated obliquely. Wings reduced to a small scale that can be perceived beneath the tegmina. Abdomen reddish brown to blackish brown, darkened toward the posterior end, cerci blackish brown. Tegument surface punctate, shiny. Total body length without cerci: ♂♂: 7.00– 10.06 mm (mean = 8.77 mm; n = 16); ♀♀: 8.25–10.62 mm (mean = 9.10; n = 18). Length of cerci: ♂♂: 2.13–4.00 mm (mean = 2.47; n = 16); ♀♀: 1.83–2.10 mm (mean = 1.95; n = 18) ( Table 3 View Table 3 ).
Etymology
The name ‘ cantabrica ’ is a feminine gender Latin adjective derived from ‘ cantabricus ’ that means belonging to or related to the Cantabrian Sea or the land adjacent to that sea, reason for which the mountain range parallel to this sea is called Cantabrian Mountains, in Spanish ‘Cordillera Cantábrica’.
Type material
Holotype SPAIN • ♂; “España, Castilla y León: León: Torrestío , Puerto de La Farrapona , 43°02′51″ N, 6°05′18″ W, 1651 m, 17-IX-2017, A.D. Cuesta-Segura leg. // MNCN_Ent 167418 // Holotipo Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & García-París , des. 2022” [red label, printed]; MNCN_Ent 167418 ( Figs 6–7 View Fig View Fig ). GoogleMaps
Paratypes
All carrying a red or white label printed (if preserved in ethanol) with “Paratipo Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & Gª-París, 2022”, from the following localities:
(1) Dry mounted specimens
SPAIN – Asturias • 18 ♂♂, 12 ♀♀; Valle del Lago, Somiedo ; 1565 m a.s.l.; 26–30 Jul. 1949; Exp. Inst. Esp. Entomología leg.; MNCN_Ent 123702 , 123706 to 123716 , 123718 to 123722 , 123724 , 123726 to 123737 • 1 ♂; same collection data as for preceding; 4–12 Aug. 1949; MNCN_Ent 123717 • 2 ♂♂, 2 ♀♀; same collection data as for preceding; 20–30 Jul. 1950; MNCN_Ent 123703 , 123725 , 123738 , 123739 • 2 ♂♂, 2 ♀♀; same collection data as for preceding; 1–10 Aug. 1950; MNCN_Ent 123704 , 123705 , 123741 , 123742 • 1 ♂; Puerto de Leitariegos ; 1575 m a.s.l.; 1–10 Aug. 1950; Exp. Inst. Esp. Entomología leg.; MNCN_Ent 123723 • 2 ♀♀; same collection data as for preceding; 1675 m a.s.l.; 1–10 Aug. 1950; Exp. Inst. Esp. Entomología leg.; MNCN_Ent 123740 , 123743 • 8 ♂♂, 9 ♀♀; Puerto de Ventana ; 43°3′31.6″ N, 6°0′10.02″ W; 1603 m a.s.l.; 19 Aug. 2017; ADC-S leg.; MNCN_Ent 167422 , 167424 , 167428 , 167429 , 167438 to 167441 , 167446 , 167455 , 167468 , 167469 , 167478 , 167499 to 167501 , 167503 GoogleMaps . – Castilla y León: León • 1 ♂; Puerto de Vegarada ; 43°2′17.08″ N, 5°28′42.39″ W; 1560 m a.s.l.; 1–23 Aug. 2008; ADC-S leg.; in pitfall trap; MNCN_Ent 167420 GoogleMaps • 2 ♂♂; same collection data as for preceding; 43°01′54.95″ N, 5°28′34.17″ W; 1684 m a.s.l.; 15 Jul. 2021; ADC-S leg.; ADC-S col. Derm_0005 , 0006 GoogleMaps • 1 ♂, 5 ♀♀; Puerto de Ventana ; 43°03′13.5″ N, 6°0′31.6″ W; 1559 m a.s.l.; 19 Aug. 2017; ADC-S leg.; MNCN_Ent 167431 , 167434 , 167442 , 167444 , 167447 , 167498 GoogleMaps • 4 ♂♂, 8 ♀♀; same collection data as for preceding; 43°03′11.3″ N, 6°00′09.6″ W; 1543 m a.s.l.; 20 Aug. 2017; ADC-S leg.; MNCN_Ent 167421 , 167430 , 167432 , 167433 , 167435 to 167437 , 167445 , 167448 , 167475 , 167479 , 167497 GoogleMaps • 2 ♂♂, 1 ♀; same collection data as for preceding; 43°03′26.3″ N, 6°00′11.7″ W; 1590 m a.s.l.; 20 Aug. 2017; ADC-S leg.; MNCN_Ent 167443 , 167476 , 167477 GoogleMaps • 19 ♂♂, 19 ♀♀; Torrestío, Puerto de La Farrapona ; 43°2′51.99″ N, 6°5′18.10″ W; 1651 m a.s.l.; 17 Sep. 2017; ADC-S and M. Sánchez Gutiérrez leg.; MNCN_Ent 167419 , 167423 , 167425 to 167427 , 167456 to 167467 , 167470 to 167474 , 167481 to 167483 , 167485 to 167496 , 167502 GoogleMaps • 3 ♂♂, 2 ♀♀; same collection data as for preceding; ADC-S col. Derm_0001 to Derm_0004 , Derm_0007 GoogleMaps .
(2) Ethanol preserved specimens
SPAIN – Castilla y León: León • 5 ♂♂; Puerto de Vegarada ; 43°01′54.95″ N, 5°28′34.17″ W; 1684 m a.s.l.; 22 Aug. 2017; ADC-S leg.; MNCN_Ent 331219 to 331223 GoogleMaps • 1 ♂, 3 ♀♀, 3 nymphs; Puerto de Ventana ; 43°03′27″ N, 6°00′15″ W; 1588 m a.s.l.; 15 Sep. 2011; M. García-París and P. Pavón leg.; MNCN_Ent 331179 to 331185 GoogleMaps • 2 ♂♂, 2 ♀♀, 3 nymphs; same collection data as for preceding; 16 Sep. 2011; M. García-París and P. Pavón leg.; MNCN_Ent 331172 to 331178 GoogleMaps .
Description
Male (holotype, MNCN_Ent 167418)
GENERAL ASPECT AND COLORATION. Male specimen, large, robust with the maximum width at posterior third of body ( Figs 6–7 View Fig View Fig ). General coloration oflive specimen reddish brown; head lighter, almost orange; pronotum, tegminae, legs and first abdominal segments, also lighter, dark-orange to honey-yellow; pronotum and tegminae somewhat transparent, not homogeneously colored because of the subjacent structures; eyes deep-black, antennae brownish, densely covered by short decumbent setae that provides gray hue to general appearance, specially to central and apical tergites; abdominal segments chestnutbrown with anterior portion of each segment lighter, dark-red; cerci chestnut-brown colored darkened toward basis ( Fig. 7 View Fig ).
MEASUREMENTS. Total body length without cerci of 9.38 mm. Cerci length 4.00 mm ( Table 3 View Table 3 ).
HEAD. Transverse (1.97 mm maximum width; 1.48 mm length without clypeus), about 0.75 times as long as wide ( Fig. 6 View Fig ). Postfrontal and coronal sutures fine but distinct, postfrontal sutures shorter than distance between them and eyes. Two deep transverse short incisions internal to eyes. Frons flat, slightly darkened. Posterior margin of the head feebly emarginated at middle. Eyes black, almost triangularly shaped, length approximately 0.5 times that of head behind eyes. Antennae with 12 antennomeres (right one missing two); antennomere I long (0.48 mm), narrowed basally, widened terminally, almost equal to antennomeres II–III combined (I/II+III = 0.97); antennomere II short (0.14 mm), but longer than wider; antennomere III (0.34 mm) longer than antennomere IV (0.26 mm), slightly widened terminally. Antennomeres II–III cylindrical, all others sub-conical. All antennomeres pubescent; dense, fine and white pubescence from middle of third antennomere to apical end. Maxillary palpi with articles I and II relatively long but shorter than third, sub-cylindrical and slightly widened distally. Article III subellipsoidal, elongated, with very thin base.
PRONOTUM. Smooth, subquadrate (1.67 mm long, 2.04 wide; 1.22 times as wide as long), sides slightly widened posteriorly ( Fig. 6 View Fig ). Anterior margin nearly straight, anterior angles with well-marked ridge that continues along lateral margin, sometimes less clearly marked. Posterior margin rounded. Median sulcus fine but distinct up to half of pronotum. Mesonotum and metanotum not visible. Sternal plates typical for genus. Tegmina short, truncated, with inner edge shorter than outer, clearly wider than pronotum and wider at end than at base. Wings reduced to small scale visible under tegmina.
LEGS. With femora and tibiae compressed. Profemora much more swollen meso- and metafemora, with more than 15 strong setae on inside of anterior side, a few setae on mesofemora and none on metafemora. Tibiae covered with thick and fine golden setae. Three tarsal segments, metatarsomere I long and subcylindrical, longer than pro- and mesotarsomeres, II spatulated, incorporating insertion of third, III elongated widened distally accommodating two strong curved claws, directed ventrally. Length of metatarsomere I (0.78 mm) almost equal to metatarsomeres II and III combined (0.76 mm; I/II+III = 0.94).
ABDOMEN. Stout, relatively long and depressed, slightly widened in posterior third; fine and densely punctate; tergites convex ( Fig. 6 View Fig ). Lateral glandular folds feeble-marked almost as scars, well visible on tergites 3–4. Last tergite transverse; surface partially rugose and punctuated. Penultimate sternite with semicircular posterior margin, with slightly prominent apex. Last tergite with slightly concave sides; slightly broadened posteriorly. Posterior margin of last tergite rounded and very slightly elevated. Pygidium completely visible, very wide, rectangular. Cerci long, sinuous and curved, with two teeth very apparent. First one (dorsal tooth) at base of cerci, in dorsal position and of triangular shape, dorsointernally directed and with apex pointing forward; visible in dorsal and lateral views. Second one (internal tooth) is at approximately ³/5 of the length of cerci, in ventro-internal position, laminar and triangle-shaped with broad base, visible in dorsal and ventral views.
GENITALIA. Robust (described from large number of paratypes, Fig. 8A View Fig ). Metaparameres fully developed, short, robust, straight and slightly narrowed at apex and with obtuse or rounded tips slightly directed inwards. Virga almost straight, slightly sinuous and short, shorter than metaparameres. Basal vesicle relatively small, kidney-shaped, strongly curved, with small sclerotized plate lacking supplementary sac associated to basal vesicle (present in smaller specimens) ( Fig. 8A View Fig ).
Variability in males Male cerci and last tergite extremely variable, but most specimens can be classified in four different phenotypes (I, II, III, and IV) ( Figs 3–4 View Fig View Fig , 7 View Fig ). In addition to these phenotypes, there are also specimens with intermediate characteristics (I–II, II–III, and III–IV) ( Figs 3–4 View Fig View Fig ).
Phenotype IV: as described in the holotype.
Phenotype III: last tergite with slightly concave sides; slightly broadened posteriorly; posterior margin with central protuberance projected upwards and rough at its edge; with the posterior side slightly inclined forward and visible in dorsal view, leaving the pygidium completely visible. Pygidium very wide, rectangular. Cerci symmetrical, broad at the base and separated from each other, medium size (2.81 mm) and curved, with a digitiform tooth at the inner edge located approximately ⅓ of its length, which is initially directed inwards and then upwards. In lateral view, these teeth do not protrude or protrude very slightly from the upper part of the cerci, which are slightly convex. The cerci in their final ⅔ are narrower, being of almost round section, except for a flat area located interno-ventrally clearly visible in ventral view.
Phenotype II: last tergite with parallel sides, with a median longitudinal furrow not very marked but visible; posterior margin with a triangular central protuberance projected backwards that partially or totally covers the pygidium. Pygidium narrow. Cerci similar to phenotype III, except for their smaller size (2.24 mm) and by the more robust digitiform tooth. In lateral view, these teeth protrude slightly but clearly from the upper part of the cerci.
Phenotype I: last tergite with parallel sides, with a median longitudinal furrow not very marked but visible; posterior margin with a central protuberance emarginated, paralleled to the body axis; this protuberance covers partially or totally the pygidium. Pygidium narrow. Cerci symmetrical, long (2.37 mm), broad at the base but clearly separated from each other, short and slightly curved, thinner and longer than in phenotype II; with a robust triangular tooth at the inner edge located near to the base and tilted up about 45 degrees. In lateral view, cerci are practically straight; the inner teeth do not protrude or protrude very slightly in lateral view. The cerci in their apical ⅔ are tapering, with almost round section, except for a flat section area located interno-ventrally visible in ventral view.
Males of P. cantabrica sp. nov. have fully developed metaparameres, slightly narrowed at the apex and with obtuse or rounded tips ( Fig. 8 View Fig ). However, just as there is variability in the morphology of male cerci, there is also intraspecific variability in male genitalia, including the size of the parameres and the basal vesicle shape. Small specimens present a supplementary sac associated to the basal vesicle, which is absent in large specimens (phenotype III–IV). The latter (phenotype III–IV) present a small sclerotized plate on the basal vesicle.
Female
Similar to male in most characters except last tergite with slightly concave sides, clearly narrowed posteriorly; with a median longitudinal furrow. Two very small protuberances on the posterior margin, just above the insertion of the cerci. Female cerci have simple and straight contiguous branches, short, wide at the base and slightly curved inwards just at the apex ( Fig. 5A–B View Fig ).
Eggs
Eggs ivory yellow ( Fig. 5D View Fig ). Some may be somewhat deformed, presenting small flat areas by contact with other eggs. The egg masses remain grouped. Eleven eggs laid in captivity by one female from Puerto de La Farrapona were measured (average, minimum-maximum measures): length (mean = 1.15 mm, range = 1.13–1.17 mm, n = 11), width (mean = 0.94 mm, range = 0.92–0.95 mm, n = 11) and a length/width ratio (mean = 1.22 mm, range = 1.18–1.27 mm, n = 11).
Nymphs
A female from Puerto de Vegarada with 34 1 st instar nymphs was held for study. As the nymphal stages aged, the number of antennomeres, the width of the head and the length of the body and cerci increase ( Table 4 View Table 4 ). Width of head seems reliable to determine nymphal stage, since ranges across stages do not overlap. The rest of the characteristics are common for the different nymphal stages ( Fig. 5C View Fig ). 1 st and 2 nd instar: head and dorsum of body dark. Legs almost transparent except for femora with central ring and base of tibiae slightly darkened. Dark antennomeres, pale at both ends. Postfrontal sutures reaching eyes and coronal suture extending to first abdominal segment, this included. Cerci with very fine granulation on inner edge, only visible under stereo microscope. From 3 rd instar on: body completely dark, except light legs. Darkened ring of femora generally wider, occupying almost basal half. Anterior half of tibiae darkened. Cerci with marked granulation on interior edge.
Comparisons
Pseudochelidura cantabrica sp. nov. differs morphologically from all other species of the genus ( P. sinuata , P. galvagnii and P. orsinii ) and also genetically from the more similar species, P. sinuata .
Male cerci are extremely variable in P. cantabrica sp. nov. and P. sinuata ( Figs 2–4 View Fig View Fig View Fig , 9 View Fig ), with marked differences in the shape of the cerci of the larger males: phenotypes III–IV ( Figs 2E View Fig , 3E View Fig ) and IV ( Figs 2F View Fig , 3F View Fig ), but not so clear across other phenotypes ( Cuesta-Segura 2012). In the phenotype III–IV, the internal tooth located at ⅓ of the cerci is almost non-existent in P. cantabrica , while it is very distinct, flat, with a curved anterior edge and a straight posterior edge in P. sinuata . In addition, P. cantabrica has slightly marked internal teeth, located at ½ of the cerci, which do not exist in P. sinuata . Males of P. sinuata phenotype IV have longer cerci and a more marked dorsal tooth at its base than in P. cantabrica . The internal teeth in P. sinuata are at ¼ of the cerci, triangle-shaped with a narrow base, pointing posteriorly, and are visible in dorsal, ventral and lateral views. However, the internal teeth of P. cantabrica are at approximately ³/5 of the cerci, triangle-shaped with a very wide base, pointing inwards and visible only in dorsal and ventral views. Differences in cerci between males of phenotypes I, II and III are subtle, involving teeth shape and direction (see Figs 2–3 View Fig View Fig ). Females are so far indistinguishable.
Steinmann (1979, 1981) provided an illustration of the male genitalia of three phenotypes (II, III and IV) of P. sinuata (sub P. sinuata , P. montuosa and P. minor ) from the Pyrenees. The genitalia of male specimens from the Pyrenees (MNCN_Ent 123583, 123639, 123640) match the description presented by Steinmann (1979, 1981) and Albouy & Caussanel (1990). Male genitalia of P. sinuata and P. cantabrica sp. nov. do not differ significantly. Both species have metaparameres fully developed, slightly narrowed at the apex and with obtuse or rounded tips. The virga is relatively short and thick and is located within the genital lobe, with a basal vesicle of the “forficuloid” type ( Steinmann 1979). Our study supports Steinmann’s (1979, 1981) descriptions of phenotypes II (sub P. minor ) and III (sub P. montuosa ) differing from phenotype IV by showing a supplementary sac associated to the basal vesicle. Small specimens of P. cantabrica also present a supplementary sac associated to the basal vesicle. In addition, phenotype IV of P. sinuata and P. cantabrica presents a small sclerotized plate on the basal vesicle.
Male cerci of P. galvagnii and P. orsinii show limited morphological diversity when compared with those of P. cantabrica sp. nov. and P. sinuata . Vigna-Taglianti (1999: figs 7–9) reported two main phenotypes in P. galvagnii . A first one with large curved cerci, very similar to those of P. cantabrica ( Fig. 3F View Fig ) with a dorsal tooth near the base and a long laminar ventrolateral internal projection and a second one with long, thinner cerci, slightly curved, almost straight, with a reduced dorsal tooth and a poorly developed ventrolateral internal projection. This second phenotype (Vigna-Taglianti 1999: fig. 9) does not show correspondence in P. cantabrica , where all males present markedly curved cerci. Large curved cerci of the first phenotype in P. galvagnii (Vigna-Taglianti 1999: figs 7–8) present a series of internal denticles at the dorsal projection level, that are totally absent in P. cantabrica . Male virga of P. galvagnii is markedly arcuate distally (Vigna-Taglianti 1999: fig. 11). Male cerci of P. orsinii are very different from those of P. cantabrica . Male cerci of P. orsinii present the lateral inner projection strongly directed inwards resulting in a double concavity that make cerci appearing ‘3’ shaped, even those of specimens with smaller cerci (Vigna-Taglianti 1999: figs 2–4). As in the case of P. galvagnii , the phenotype of P. orsinii with smaller cerci is not similar to those in P. cantabrica , where all males present markedly curved cerci, and not almost straight. Male virga of P. orsinii is slighltly arquate distally ( Maccagno 1933: fig. 11; Vigna-Taglianti 1999: fig. 6) while it is relatively short and thick in P. cantabrica .
Differences between P. galvagnii and P. orsinii are basically at the same level as those reported here between P. sinuata and P. cantabrica sp. nov., but intraspecific diversity is much limited in P. galvagnii and P. orsinii (Vigna-Taglianti 1999) .
Geographic range
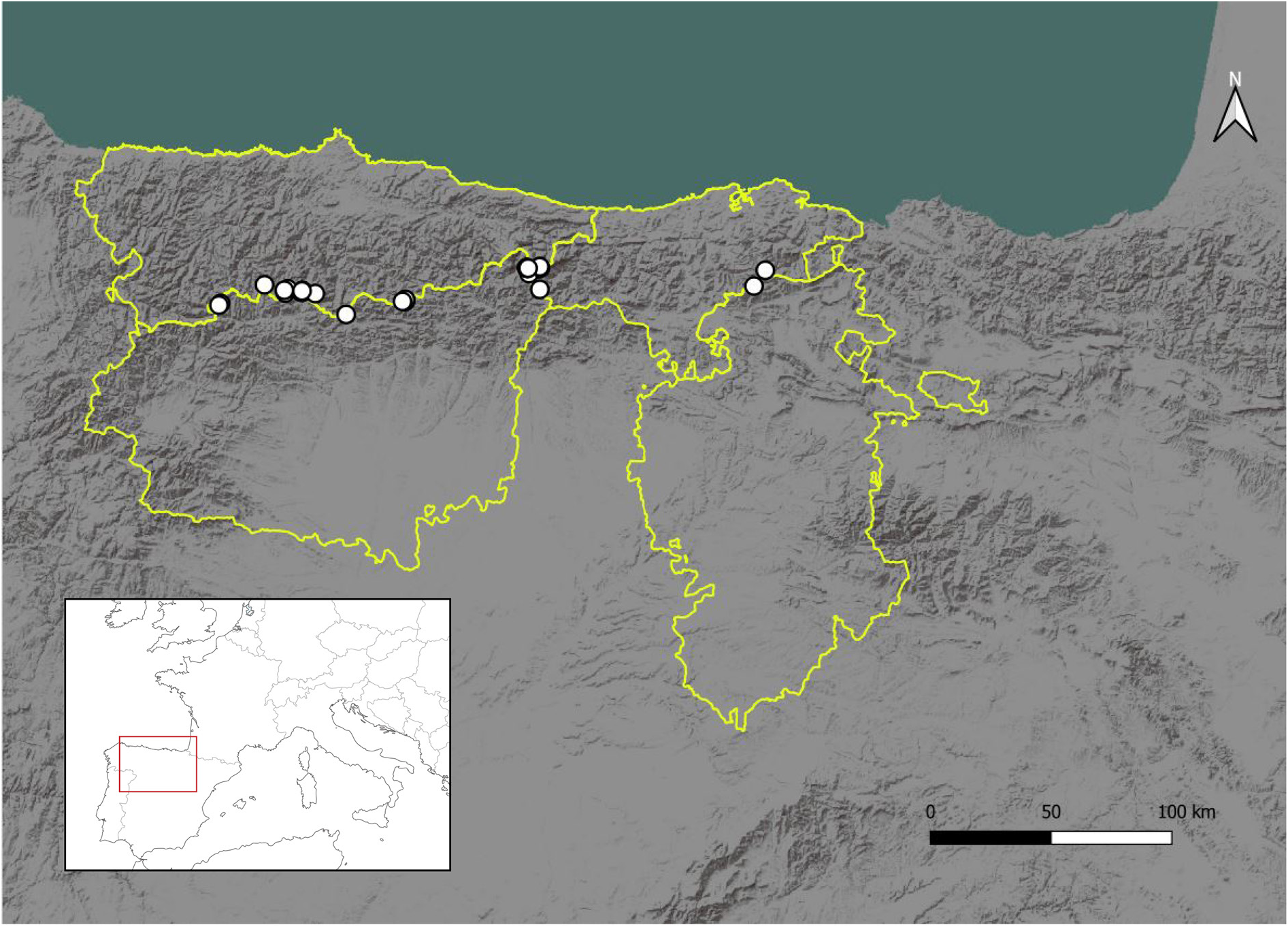
Pseudochelidura cantabrica sp. nov. occurs along to the Cantabrian Mountains ( Fig. 10 View Fig ). At the moment, the species has been located in the Spanish provinces of Asturias, Burgos, Cantabria, and León, at altitudes between 1128 and 2184 m.
In addition to the localities included in the type series and in the material and methods section (see above), P. cantabrica sp. nov. has previously been reported from diverse localities under different names (see also comments at the end of the synonymy list). According to the illustrations provided by Harz & Kaltenbach (1976), the records of P. sinuata from Peña Vieja and Asturias (Picos de Europa, Spain) correspond to P. cantabrica . Most of the specimens in the MNCN Collection mentioned by Lapeira & Pascual (1980) sub P. sinuata from the Cantabrian Mountains: Peña Vieja (Picos de Europa, Spain), Valle del Lago and Leitariegos (Asturias, Spain), are considered paratypes of P. cantabrica in the present work, except the specimens from “Sorbea [Gorbea], Bilbao” and “Torrebarrio, León” which correspond to female specimens of Forficula (possibly F. lesnei Finot, 1887 ). We have not found the two specimens mentioned by Lapeira & Pascual (1980) sub P. sinuata with the labels “Ladera del Coriscao, Santander, 21-VII-54 (Peris)” and “W. from Lagoa Comprida, Serra Estrella, Portugal, 28-VIII-42 (Machado)”. Bivar de Sousa (1997) commented that the specimen from Lagoa Comprida corresponded to a nymph of Mesochelidura bolivari , and that consequently there were no confirmed records of Pseudochelidura in Portugal. On the contrary, because of the proximity of Ladera del Coriscao to known localities of Pseudochelidura , we consider that this specimen likely corresponds to P. cantabrica . According to the description provided by Neira Artidiello (2009), the male of P. sinuata captured in the Macizo de las Ubiñas (Asturias, Spain) corresponds to P. cantabrica . The specimens from Torrestío and Redipuertas (León, Spain) cited as P. minor by Cuesta-Segura (2012) and studied in the present work, also correspond to P. cantabrica . Finally, the male from Fuente Dé (Cantabria, Spain) cited as P. sinuata by Pérez-Valcárcel et al. (2022) also corresponds to P. cantabrica .
Notes on natural history
Pseudochelidura cantabrica sp. nov. is usually found in mountain meadows and pastures ( Fig. 11 View Fig ). These mountain pastures are usually grazed by transhumant cattle during the summer, usually cows, sheep and horses. In a biodiversity study using pitfall traps at the mountain pass of Vegarada in the summers of 2004 and 2008, specimens of P. cantabrica were found only in the pastures, but not under the bushes (mixed Calluna Salisb. and Vaccinium L.) ( Cuesta-Segura 2012, 2016 sub P. minor ).
Pseudochelidura cantabrica sp. nov. is an uncommon species occasionally found under rocks or dry cow dung and can locally be abundant but populations are not continuous, patchily distributed and difficult to locate. A study surveying 15 mountain passes along the Western Cantabrian range, by using pitfall traps, only detected the presence of the species at a single locality ( Cuesta-Segura 2012 sub P. minor ). Populations of P. cantabrica coexist with the ubiquitous Forficula dentata Fabricius, 1775 . Abundance percentages of one or the other species vary across zones, P. cantabrica being less abundant in most cases ( Table 5 View Table 5 ). Both species, but especially P. cantabrica , were more abundant in locations facing north ( Table 5 View Table 5 , zones 2 and 8).
During the summer of 2017, females were more frequent than males ( Table 5 View Table 5 ). Males with a diverse cerci morphology (phenotypes I, II, III and IV) and females (one of which with nymphs) were observed together in the Vegarada mountain pass under a single cow pat. In general, phenotypes I and II were the most abundant, III and IV being scarce. As an example, at Puerto de La Farrapona, 17 Sep. 2017, we observed (only a few specimens collected), 95 nymphs, 46 females and 33 males; 23 males (70%) corresponding phenotype I, four (12%) to phenotype II, one (3%) to phenotype III, one (3%) to phenotype IV (the holotype), and four (12%) to intermediate phenotypes (I–II, II–III, and III–IV). One of the females kept alive from Puerto de La Farrapona laid 27 eggs and while initially she took care of them, about two weeks later she ate them. The same female laid 17 eggs three weeks later, and ate them too.
Synonymy list and nomenclatural combinations of Iberian Pseudochelidura Verhoeff, 1902
Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & García-París sp. nov.
Pseudochelidura cantabrica Cuesta-Segura, Jurado-Angulo & García-París sp. nov. Terra typica: “ España: Castilla y León: León: Torrestío, Puerto de La Farrapona, 43°02′51″ N, 6°05′18″ W, 1651 m ”.









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |