
Salvia albiterrarum J.G.González J.G.González & Art.Castro, 2013
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.93.2.1 |
|
persistent identifier |
https://treatment.plazi.org/id/890D87BC-7A7C-FFDA-8F9C-7C97B1A2F866 |
|
treatment provided by |
Felipe |
|
scientific name |
Salvia albiterrarum J.G.González J.G.González & Art.Castro |
| status |
sp. nov. |
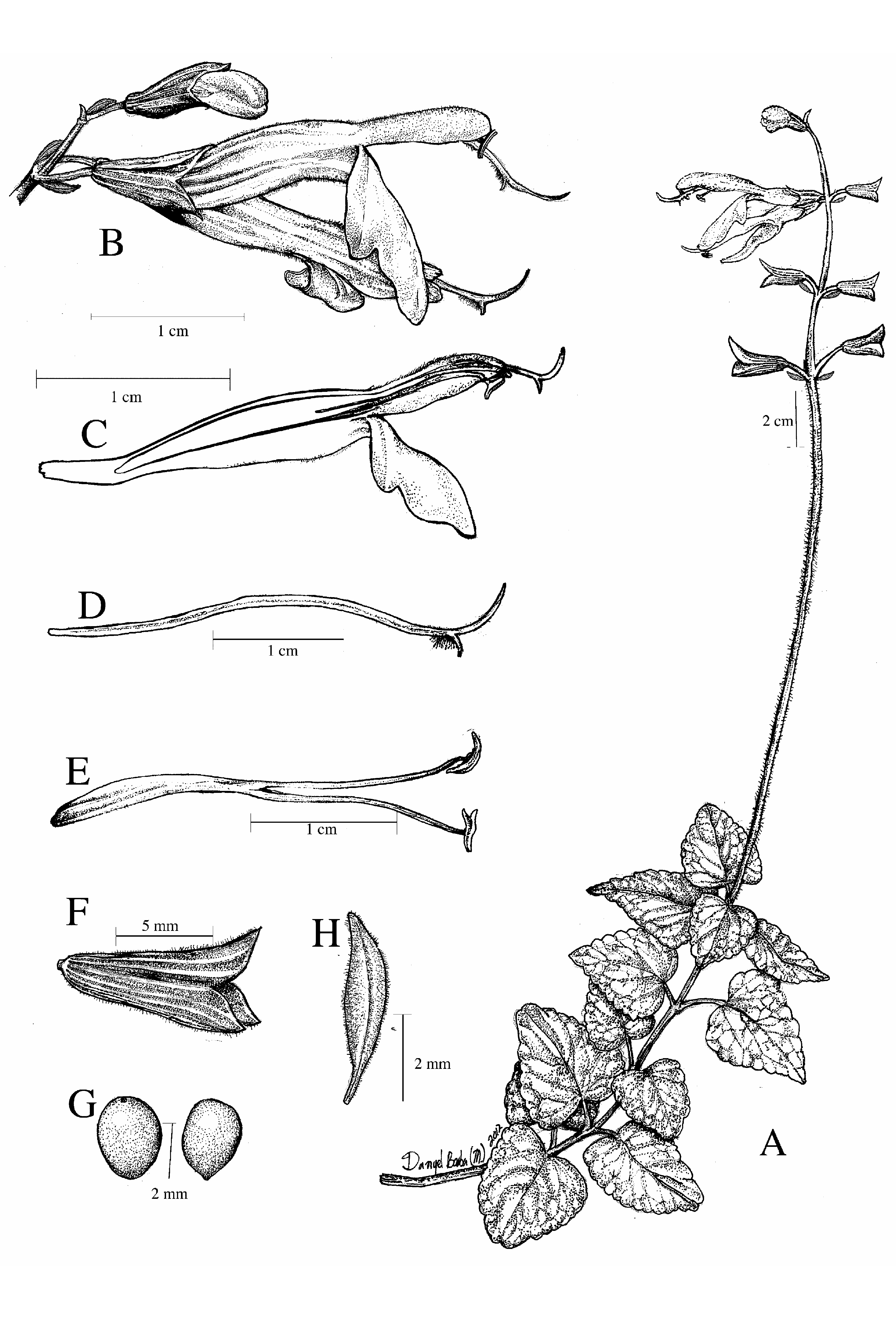
Salvia albiterrarum J.G.González J.G.González & Art.Castro View in CoL , sp. nov. ( Figs. 1 E–F View FIGURE 1 , 2 B View FIGURE 2 , 3 View FIGURE 3 , 4 C View FIGURE 4 , and 5 View FIGURE 5 )
A Salviae puganae corollarum tubis longioribus (17.3)18.6–23.5 vs. 8.7–9.6(–12.7) mm), signis nectariferis nullis vel reductis, filis longioribus (6.8–8.6 vs. 3.8–5.2 mm), gubernaculis longioribus (16.7)20.0–28.5 vs. 6.3–11.2 mm) et sine dentibus, stylis longioribus (26.4–37.0 vs. 13.0–15.0 mm) et longe exsertis, ramis stigmatiferis inferioribus rectis (vs. sigmoideis vel arcuatis) statim dignoscenda.
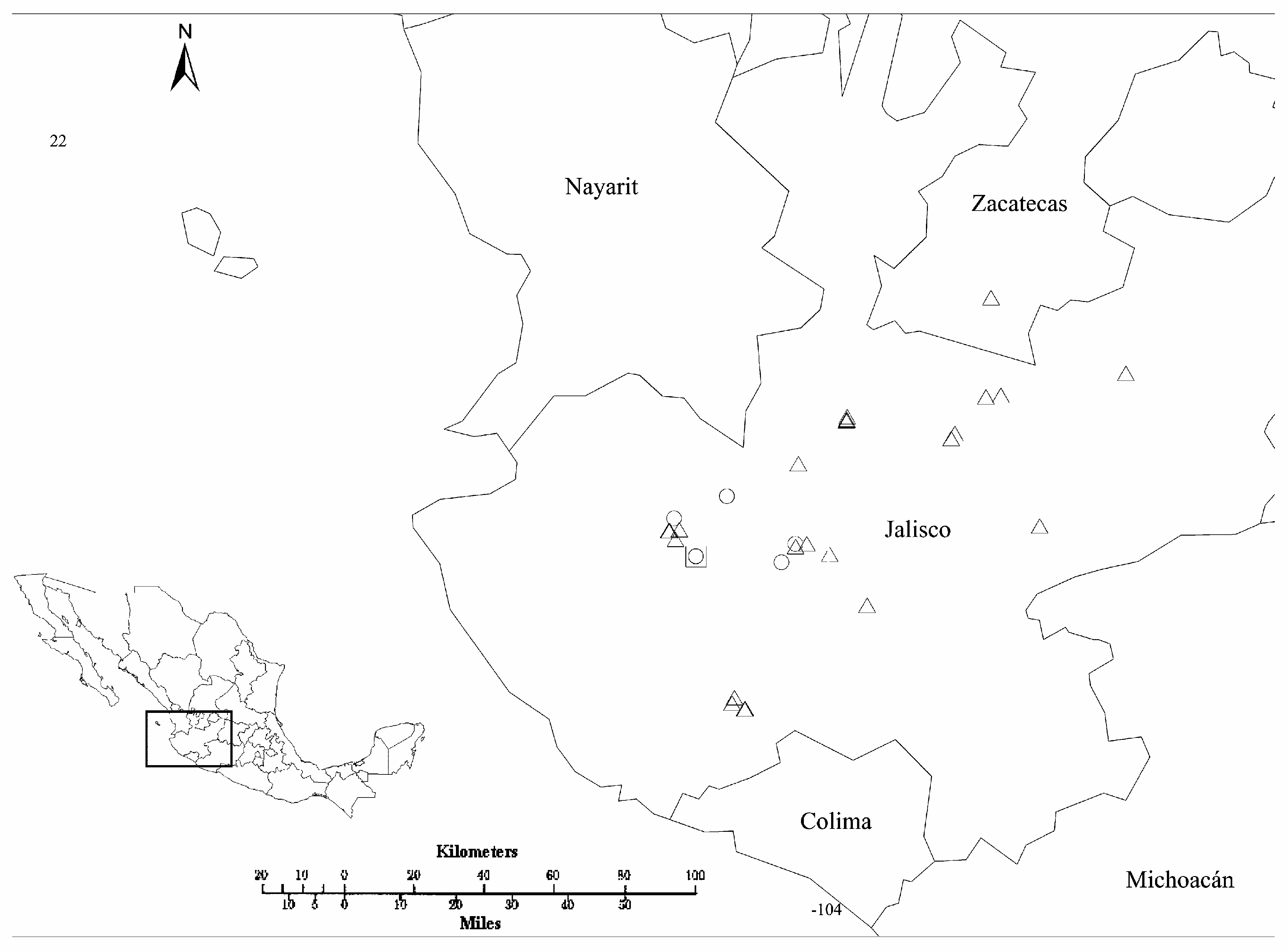
Type: — MEXICO. Jalisco: Cuautla: 12.8 km al SE de Los Volcanes por la carretera rumbo a Cuautla, entre el crucero de Fresno Hueco y Tierras Blancas, 20.249ºN, 104.484ºW, 1967 m, 31 July 2012 (fl, fr), J. González-Gallegos, A. Castro-Castro y M. A. Carrasco-Ortiz 1246 (holotype IBUG!, isotypes CIIDIR!, CIMI!, ENCB!, GUADA!, HUAA!, HUMO!, IEB!, MEXU!, OAX!, SERO!, UAGC!, XAL!, ZEA!) GoogleMaps .
Perennial herb, erect, 25–60 cm tall, stems sparsely pilose to glabrous. Leaves with petioles (1.0–) 1.8–4.6 cm long, sparsely pilose; blade ovate-deltoid, 2.4–5.7 × 1.8–5.2 cm, acute to acuminate, obtuse to rounded at apex, truncate to cordate or sometimes oblique at base, broad serrate to crenate margin, surface bullate and lustrous above, paler beneath and covered with glandular dots, both pilose on the veins, otherwise glabrous. Inflorescence terminal, 19–40 cm long, with 4–15 verticillasters, these 2-flowered, or rarely up to 6-flowered, the lowermost 2.0– 6.5 cm apart from each other toward the base, floral axis hispidulous and covered with short glandular-capitate hairs between the last leaves and the first verticillaster, and sparsely pilose upward. Floral bract lanceolate to elliptic-lanceolate, 3.8–5.6 × 0.5–1.0 mm, deciduous, acute at apex, truncate at base, entire margin, outer surface pilose, inner one glabrous; bracteoles present at the base of each pedicel, linear, 1–2 × 0.1–0.2 mm, deciduous, pilose. Flower with pedicel (2.2–)3.0–4.0 mm long, up to 7.2 mm long in fruit, dense and sparsely pilose. Calyx (6.3–) 7.8–9.8 mm long, 4.0– 5.2 mm wide at the throat (up to 11 × 7.5 mm in fruit), glandular-dotted, externally pilose on the veins, internally glabrous and sometimes verrucose toward the apex, lips acute and ciliate at the margin, the upper one 7-veined and entire. Corolla magenta, color guides above the lower lip absent to scarcely developed, light magenta but not white, tube, upper lip and ventral portion of the lower one pilose; tube (17.3–) 18.6–23.5 mm long, not ventricose, 5.0– 6.8 mm wide at midportion, straight at the base, internally epapillate and without bolds; upper lip (7.2–)9.0–13.0 mm long, lower lip 9.6–14.3 × (8.2–) 9.8–12.4 mm. Stamens included; filament 6.8–8.6 mm long; connective (16.7–)20.0– 28.5 mm long, straight, not geniculate, sometimes with a tiny acute tooth at ventral midportion; theca 2.0– 3.2 mm long; staminodes present above and behind the insertion of the filament, filiform. Gynobasic horn 0.8–1.0 mm long; style 26.4–37.0 mm long, pilose at apex and before the branches, exserted 3.0– 5.3 mm plus the branches from the upper lip, the lower branch acute. Nutlet ovoid, 2.9–3.4 × 2.1–2.8 mm, uniformely dark brown, glabrous and smooth.
Distribution, habitat and phenology: — Salvia albiterrarum is known only from the municipality of Cuautla, Jalisco, in a hill at west of Tierras Blancas ( Fig. 3 View FIGURE 3 ). It inhabits oak-pine forests, from 1900–2000 m a.s.l. It shares habitat with the threes Arbutus xalapensis Kunth , Agarista mexicana , Pinus oocarpa , Quercus castanea , Q. eduardii Trel. , Q. magnoliifolia , and the shrubs and herbs Dahlia coccinea , D. pugana , Eriosema diffusum (Kunth) G.Don , Euphorbia sphaerorhiza , Phaseolus coccineus L., P. pauciflorus Sessé & Moc. ex G.Don , Salvia lavanduloides Kunth , S. platyphylla , S. prunelloides Kunth , Scutellaria dumetorum , Verbesina linearis , Cyperus sp. , Habenaria sp. , Odontotrichum sp. , and Oxalis sp. It flowers and fructifies from June to August.
Etymology: —The species epithet is the latinized name of Tierras Blancas, honoring the place where it was found and which embraces the known distribution of the taxon.
Taxonomic relationships: —If it is followed the key for the sections of Salvia subgenus Calosphace provided by Epling (1939), S. albiterrarum matches better with sect. Nobiles Epling (1939: 280) and sect. Sigmoideae , though there are some characters that disagree with sectional circunscription. Species of sect. Nobiles differ in several ways from S. albiterrarum , they are shrubs more than 1 m tall (vs. perennial herbs up to 60 cm tall), without bracteoles, their flowers are bright red without color guides above the lower lip (vs. magenta with absent to poor developed light magenta guides), corolla tubes are expanded toward the apex or rarely ventricose (vs. straight and not ventricose), lower corolla lips are as long as, or shorter than upper lips (vs. upper corolla lips shorter than the lower), and with lateral lobes poor developed to absent (vs. patent lateral lobes). Species of sect. Sigmoideae differ in having invaginated corolla tubes at base and usually internally ornate with two bolds (vs. straight and naked), geniculate connectives (vs. entire), and sigmoid lower stigmatic branches (vs. acute). However, if it is considered the presence of bracteoles additionally to floral bracts in S. albiterrarum and sect. Sigmoideae , and morphological similarity in vegetative characters between the first and S. platyphylla ( Table 1, Fig. 2 View FIGURE 2 ), it results a more supported idea to conceive S. albiterrarum as part of sect. Sigmoideae instead of sect. Nobiles. It is also noteworthy that the use of Epling's proposal should be considered provisional as there is little evidence about the monophyly most of his sections ( Standley & Williams, 1973; Jenks et al. 2008).
Salvia albiterrarum resembles S. pugana , but it differs from the latter by its longer corolla tubes [(17.3–)18.6–23.5 vs. 8.7–9.6(–12.7) mm], absence of white nectar guides on the lower lip ( Fig. 1F View FIGURE 1 ), longer filaments (6.8–8.6 vs. 3.8–5.2 mm), longer [(16.7–)20.0–28.5 vs. 6.3–11.2 mm long] and not geniculate (vs. geniculate) connectives, longer styles (26.4–37.0 vs. 13.0–15.0 mm), with branches and an apical portion of 3.0– 5.3 mm exserted from the upper lip (vs. with only the upper branch exserted), and lower stigmatic branch straight (vs. sigmoid or arquate; Table 1, Figs. 4B View FIGURE 4 vs 4C and 5). Salvia albiterrarum is known only from the municipality of Cuautla in the limits with the municipality of Atenguillo, Jalisco, where it is sympatric with S. pugana ( Fig. 3 View FIGURE 3 ).
The differences remarked between the corollas of Salvia albiterrarum and S. pugana , and even S. platyphylla ( Table 1, Figs. 1A–D View FIGURE 1 vs. 1E–F and 4A–B vs. 4C), seem to reflect the two general pollination syndromes in Salvia : melithophily and ornithophily. Salvia melithophilous corollas, in contrast with ornithophilous, are distinguished by wider and extended lower corolla lips, shorter distance between nectar position and corolla tube entrance, shorter connective arms ( Wester & Classen-Bockhoff 2007), nectar guides often found on the flower entrance, presence of distinctive ventral teeth or barriers at the connectives ( Wester & Classen-Bockhoff 2011), and pollen sacs usually hidden in the upper corolla lip (Wester & Classen- Bockhoff. 2006). This set of morphological characters is exhibited by S. pugana and S. platyphylla ( Figs. 1 A–D View FIGURE 1 , 4 A–B View FIGURE 4 ), while S. albiterrarum presents longer corolla tubes and hence a longer distance between flower entrance and nectar position, longer connective arms, nectar guides lacking or reduced and restricted to flower entrance, and teeth or outgrowths at the connectives lacking ( Figs. 1 E–F View FIGURE 1 , 4 C View FIGURE 4 ); such that it coincides with the ornithophilous syndrome described in the genus ( Wester & Classen-Bockhoff 2006a, 2006 b, 2007, 2011). Moreover, in both species the pollen sacs are hidden in the upper corolla lip suggesting and active lever mechanism ( Wester & Classen-Bockhoff 2006b), which can be confirmed introducing a mechanical pressure on the posterior portion of the connectives; however, in S. platyphylla the connectives occlude the corolla tube near flower entrance to mid-portion ( Fig. 4 A View FIGURE 4 ), whereas in S. albiterrarum the occlusion is distally located (5– 8 mm from corolla base; Fig. 4 C View FIGURE 4 ), so we can assume that lever mechanism in S. albiterrarum cannot be turn on by bees ( Wester & Classen-Bockhoff 2011). Classen-Bockhoff et al. (2004) indicates that minute changes in lever arm length, shape and orientation, and relative morphometric proportions of staminal levers and pollinators might have significant effects on flower pollinator interaction, because these changes can promote reproductive isolation between sympatric species facilitating pollen deposition on some spots of the pollinator’s bodies while excluding others. Therefore, if a phylogenetic relatedness were corroborated between these species or between S. albiterrarum and any other species from sect. Sigmoideae (which also exhibit melitophilous syndrome as S. playphylla ), it would be a good model to evaluate pollinator shift from bees to birds as a force conducting speciation within this lineage.



| J |
University of the Witwatersrand |
| A |
Harvard University - Arnold Arboretum |
| M |
Botanische Staatssammlung München |
| IBUG |
Universidad de Guadalajara |
| CIIDIR |
Instituto Politécnico Nacional |
| CIMI |
Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional (CIIDIR) IPN-Michoacán, |
| ENCB |
Universidad de Autonoma de Baja California |
| GUADA |
Universidad Autónoma de Guadalajara |
| HUAA |
Universidad Autónoma de Aquascalientes |
| HUMO |
Universidad Autónoma del Estado de Morelos |
| IEB |
Instituto de Ecología, A.C. |
| MEXU |
Universidad Nacional Autónoma de México |
| OAX |
Instituto Politécnico Nacional (CIIDIR-Oax., I.P.N.) |
| SERO |
Sociedad para el Estudio de los Recursos Bióticos de Oaxaca |
| UAGC |
Universidad Autónoma de Guerrero |
| XAL |
Instituto de Ecología, A.C. |
| ZEA |
Universidad de Guadalajara, Centro Universitario de la Costa Sur |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |