
Scales
|
publication ID |
https://doi.org/10.5252/a2011n1a1 |
|
persistent identifier |
https://treatment.plazi.org/id/886CAA78-FFE0-FFC8-FFFA-0AA9FBA5FBF6 |
|
treatment provided by |
Carolina |
|
scientific name |
Scales |
| status |
|
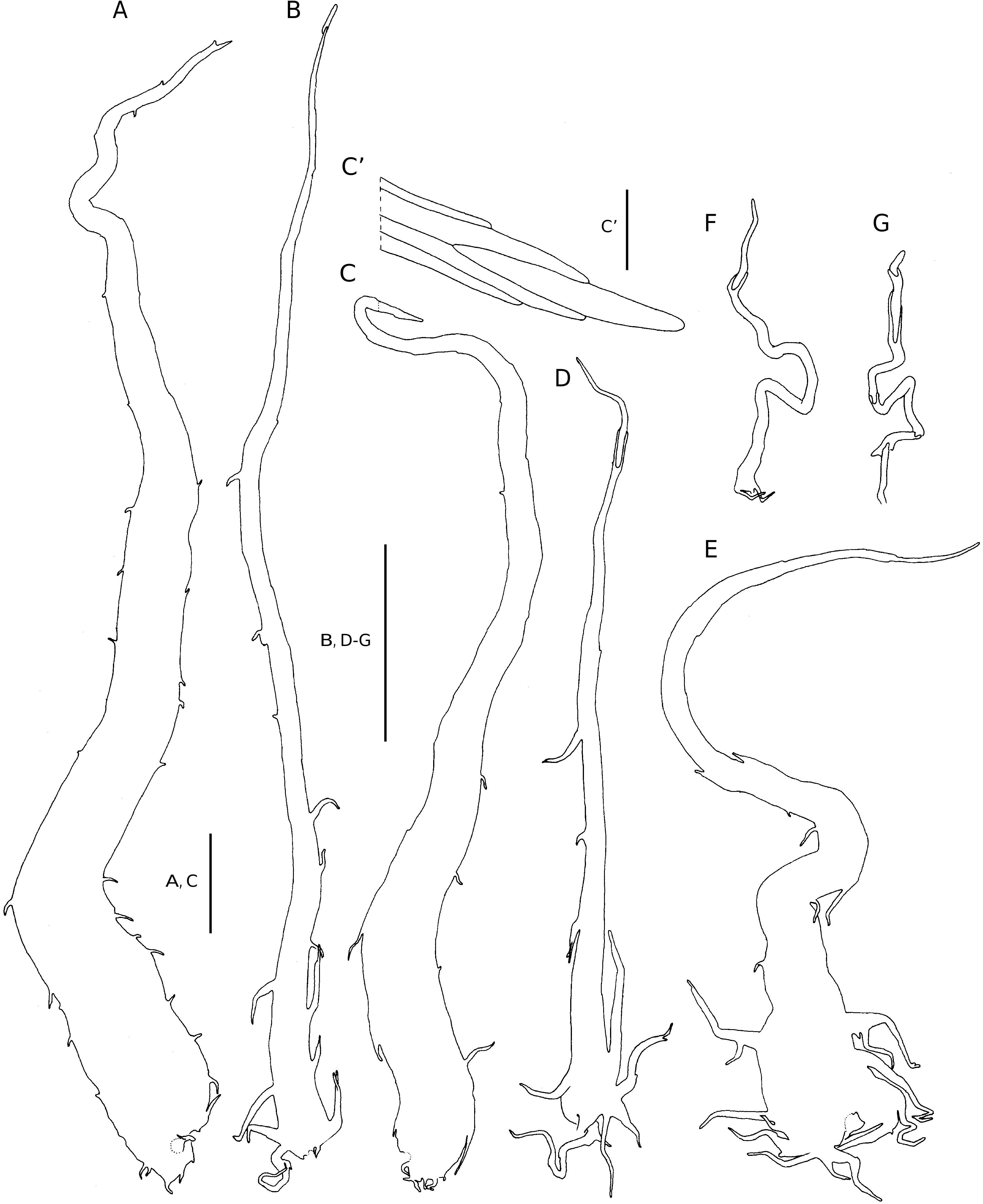
Scales are dermal appendages confined to the rhizome, frond axes and veins, but in D. wallichiana subsp. madagascariensis , small fibrils or hair-like scales occur along the lamina margin. Fibrils (= microscales [ Daigobo 1972]) are multicellular hairs and hair-like scales with sinuate joints between the cells.They often are long-stalked and bear short outgrowths near the base. These scale-like structures are confined to the lobe margins of D.wallichiana subsp. madagascariensis only ( Fig. 2F, G View FIG ). In Dryopteris and Nothoperanema , scales never occur on the lamina surfaces between the veins. In both genera, scales are mostly one cell layer thick and flat, but in a few species, some of the frond scales are often bullate, and in Nothoperanema , they are mostly patent. Scales in Dryopteris are thinwalled, membranous, or chartaceous structures. In most species, they are concolorous, and depending on their age and location, they can be stramineous, ferrugineous or castaneous. Young and immature scales are usually whitish.
Scale structure in Dryopteris shows extreme variation, but two basic types can be identified within the relevant Dryopteris species :
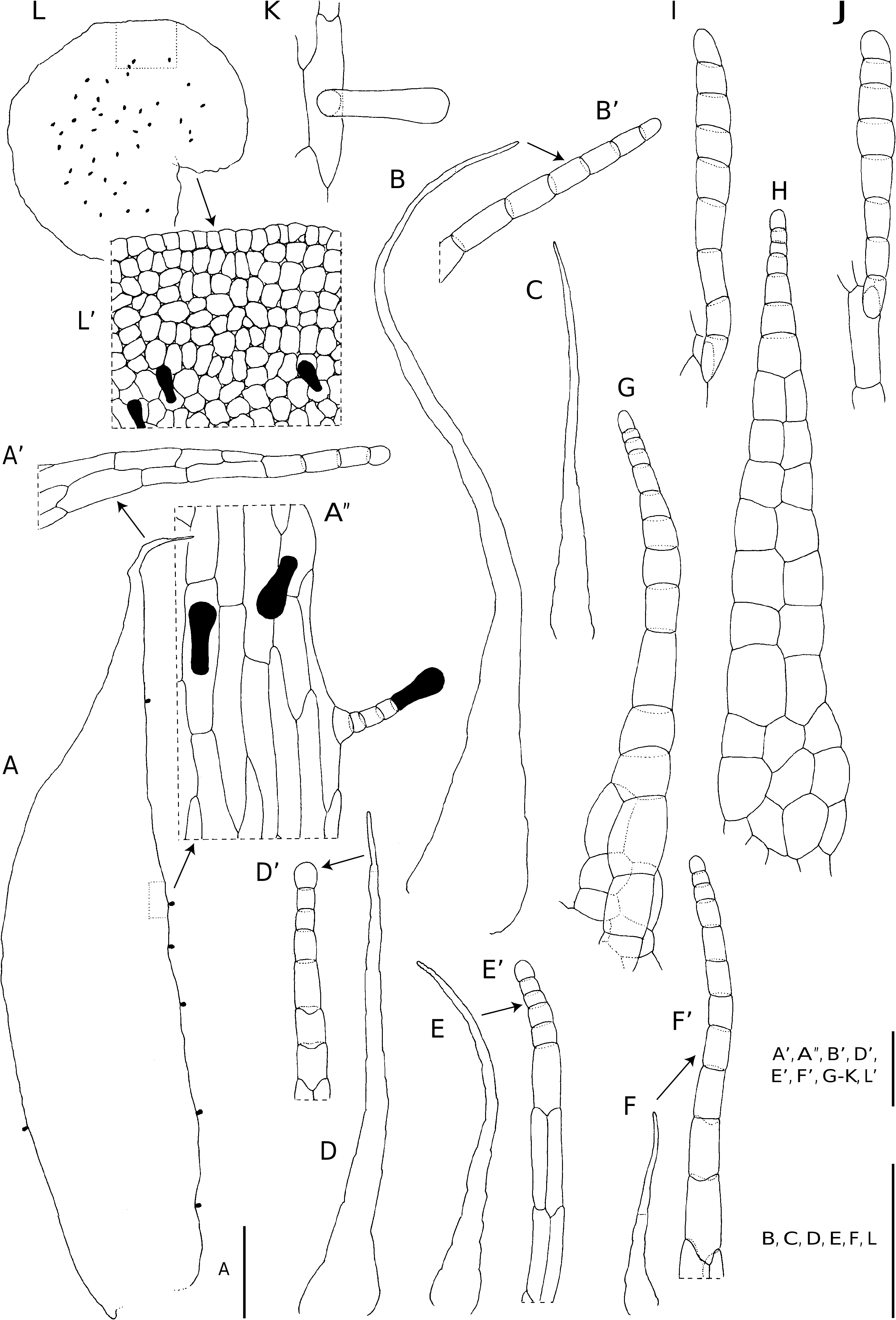
– the scales of species in sect. Fibrillosae are up to 17 mm long and 2.4 mm wide.They are chartaceous and castaneous, ferrugineous or stramineous, but the larger (stipe) scales are often bicolorous. If bicolorous, then they are ferrugineous to stramineous with (mostly central) castaneous streaks. The larger scales are subulate, broadly attached and cordate, whilst the margins are variously set with short and long, apically or basally directed, often branched outgrowths, which reduce in number and size towards scale apex. The scale apex terminates in an elliptic thin-walled cell or a subulate cell. Smaller scales are stalked, cordate to cordate-imbricate, and the margins proximally variously set with outgrowths similar to those on the rhizome scales. Short- or long-stalked filiform scales occur along the costa, veins, and lobe margins. These scales proximally bear short marginal outgrowths, but distally they are entire. The scale apex terminates in a subulate cell ( Fig. 2C View FIG ), an elliptic thin-walled cell, or the apical cell is undifferentiated. Similar scales occur in Polystichum Roth sect. Lasiopolystichum Daigobo ( Daigobo 1972) ;
– scales of the other Dryopteris sections are generally less clearly defined and they also show a greater degree of variation in overall morphology. Scales of species in these sections are basifixed and generally larger than those in sect. Fibrillosae . They are chartaceous and stramineous to ferrugineous in colour. The scales may be filiform, lanceolate, narrowly ovate or broadly ovate in outline and often bullate in sect. Purpurascentes. Pluricellular outgrowths of variable length generally occur along the scale margins, but often also on the scale laminae in D. aquilinoides . In several species capitate, clavate or oblong glands occur along the scale margins and on the laminae. The scales generally terminate in a pluricellular filiform or flagelliform apex of which the terminal cell, like the marginal outgrowths may be clavate and thin-walled, or glandular ( Figs 4 A-E; 6B, C; 15C, D; 18A, D; 20A, B, F; 26A). In D. antarctica , the marginal outgrowths and scale apex terminates in a series of moniliform cells ( Fig. 4 A-E).
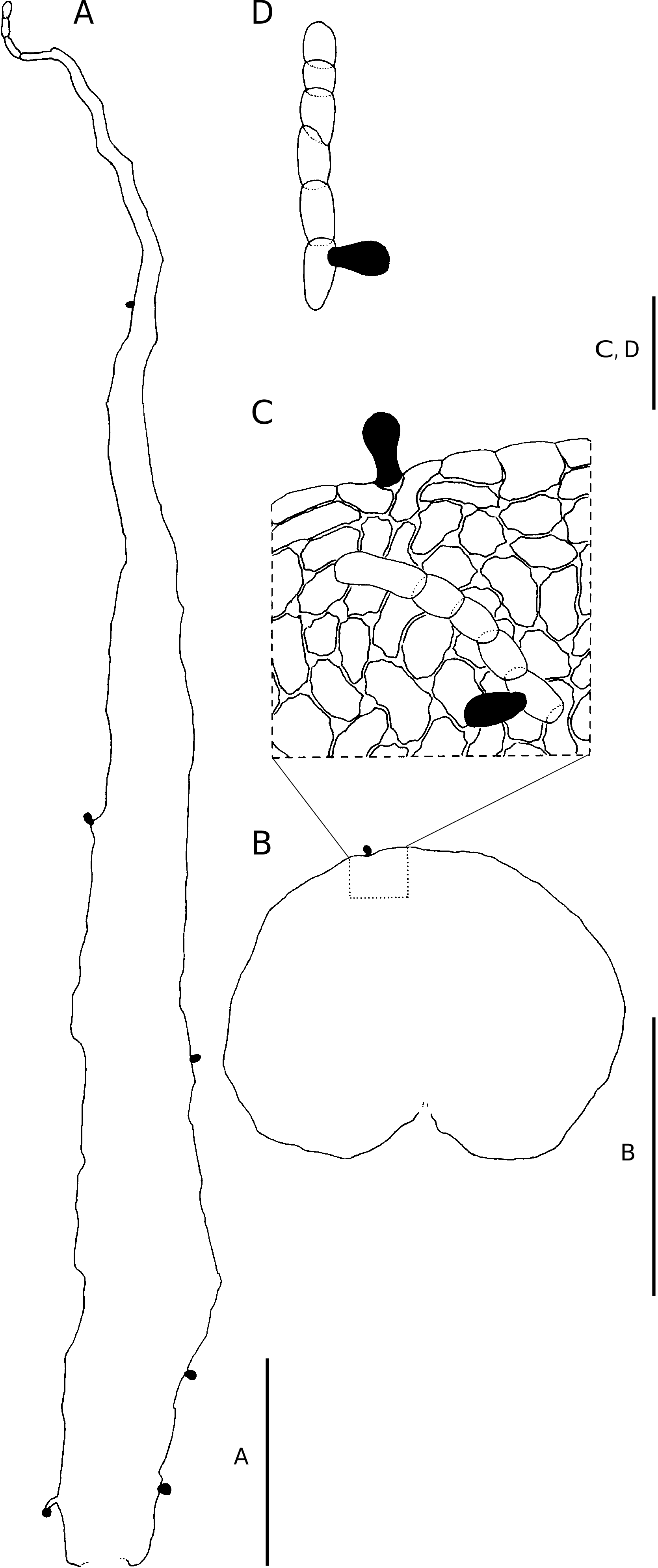
Scales in Nothoperanema are castaneous and firmly chartaceous to thinly crustaceous. In structure, they differ from any Dryopteris species in the region in that they are adnate, subulate and patent. Few-celled uniseriate hairs terminating in a clavate gland frequently occur along the scale margin with unicellular glands regularly occurring on the scale lamina margin and lamina. The scale apex terminates in a short uniseriate series of cells, the terminal cell being glandular ( Fig. 31 View FIG A’, D’, E’, F’).
SORI
Sori are borne abaxially on the lamina and are positioned medially, at or near the apex of an ultimate vein branch. Sori in the 1-pinnate-pinnatifid Dryopteris species are mostly borne on the anadromous secondary vein branches, but the bearing of sori on the catadromous secondary vein branches are not uncommon. The venation in Dryopteris species with more divided laminae generally does not show any differentiation between sterile and fertile fronds, but in D. antarctica , which belongs in sect. Lophodium , and N. squamiseta , the sori are borne at or near the apex of mostly shortened anadromous vein branches.
INDUSIUM
All the Dryopteris species in the study area, with the exception of D. manniana , are indusiate. The indusium is reniform and the wing is mostly one cell layer thick, but at the point of attachment, it is several cell layers thick. The margin is repand or entire. The anticlinal cell walls are sinuose to deeply lobed. In Nothoperanema , the transverse cell walls are straight to shallowly lobed. Indusia are persistent, but they may be lost as a result of mechanical damage. The structure is generally uniform in colour and whitish to almost translucent in live material, becoming pale to dark brown when dry. Clavate glands are often borne along the margin of some species, but in others glands also occur on the indusium lamina. The occurrence of many clavate glands on the indusium lamina is a characteristic of N. squamiseta ( Fig. 31L View FIG , L’). In addition, a few short hairs near the point of attachment may also be present on the indusium lamina. Dryopteris aquilinoides and D. wardii are the only species where uniseriate hairs occur on the indusium lamina ( Fig. 28B, C View FIG ).
SPORANGIUM
Dryopteris and Nothoperanema sporangia are biconvex in facial view and circular to slightly obovate in outline in lateral view.The annulus is vertical and interrupted by the stalk. The number of indurated annulus (or bow) cells per sporangium is variable ( Table 2), and a well-defined stomial region can be identified.
Cells of the stomial region are thin-walled. The two stomium cells themselves are distinct in being larger with slightly thicker walls. The average number of hypostomial cells per sporangium is three, but as few as two and as many as five are known. Similarly, the average number of epistomial cells per sporangium is three, but as few as one and as many as four have been observed.
The sporangium stalk is composed of three rows of cells for most of its length, but the base may be one or two cells thick. The length of the stalk and also its cellular structure vary considerably. The stalk is generally long and consists of slender cells, but it is often composed of short cupiform cells. The capsule generally breaks from the stalk through the tearing of the stalk cells, just below the capsule. Although the stalk is mostly simple, one or more clavate glands frequently occur on it, or it may bear a pluricellular isocytic hair from near the base. In D. wardii , the hairs are composed of relatively long cells, with the apical cell pyriform and gland-like.
SPORES
A general descriptive overview of the spore surface, wall structure and spore diversity in Dryopteris and Nothoperanema is provided by Tryon & Lugardon (1990). Spores of the species dealt with here are ellipsoidal, monolete, brown, and the perispore folded to form low tubercules, narrow or broad compressed reticulate ridges that are densely echinulate, minutely rugulose, minutely scabrous, ruminate, or with narrow to broad reticulate ridges.
As with stomata, a correlation appears to exist between spore size and ploidal level within a group of related taxa. It was suggested that spore size can contribute in establishing hypotheses of evolutionary relationships within related polyploid complexes ( Barrington et al. 1986). Dryopteris bojeri has the smallest spores and D. antarctica the largest ( Table 3).
The number of spores per sporangium is mostly 64, except in D. wallichiana subsp. madagascariensis , where only 32 spores per sporangium are produced, suggesting it to be an obligate apomict.
Spores of the putative hybrid are aborted.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.