
Dryopteris kilemensis (Kuhn) Kuntze
|
publication ID |
https://doi.org/ 10.5252/a2011n1a1 |
|
persistent identifier |
https://treatment.plazi.org/id/886CAA78-FFCB-FFFE-FF38-0AA9FC29FD33 |
|
treatment provided by |
Carolina |
|
scientific name |
Dryopteris kilemensis (Kuhn) Kuntze |
| status |
|
Dryopteris kilemensis (Kuhn) Kuntze View in CoL
( Figs 25 View FIG ; 26 View FIG )
Revisio generum plantarum 2: 813 (5 Nov. 1891), as “ kilmensis ”. — Aspidium kilemense Kuhn, Filices africanae: 24 (Oct. 1868), as “ kilmense ”. — Nephrodium kilemense (Kuhn) Baker , Synopsis filicum , ed. 2: 498 (Oct. 1874), as “ kilmense ”. — Type: In Kilma ad radices montis Kilimandjaro regionis Dschagga, 3-4000 ped., 1864, Kersten 46 (holo-, B20 0051628!; iso-, B20 0058467!).
Nephrodium lastii Baker, Annals View in CoL of Botany 5: 324 (1891).— Aspidium lastii (Baker) Hieron., Die Pflanzenwelt Ost-Afrikas und der Nachbargebiete View in CoL 1, C: 85 (Jun. 1895). — Dryopteris lastii (Baker) C.Chr. View in CoL , Index filicum : 274 (18 Nov. 1905). — Type: Mozambique, Namuli, Makua Country, 1887, J.T. Last s.n. (holo-, K!, 2 sheets).
Dryopteris platylepis Rosenst., Feddes Repertorium View in CoL 4: 4, 5 (1907). — Type: Africa orient. germ. ( Tanzania), Kilimanjaro, 4000 m, in humidis sylvae primaevae, 1906, P. Daubenberger 37 (holo-, M!; iso-, M!).
Dryopteris remotipinnulata Bonap. , Notes ptéridologiques 16: 178, 179 (14 Apr. 1925). — Type: Madagascar, Massif de Manongarivo , vers 1700 mètres d’altitude, bois humide, rocailles, mai 1909, H. Perrier de la Bâthie 7619 (holo-, P00349503!; iso-, P00483237!, P00483238!).
Dryopteris cordipinnula C.Chr. , Catalogue des plantes de Madagascar. Pteridophyta: 27 (Feb. 1932), nom. nud. (McNeill et al. [2006] Art. 32.1(d)); Christensen, Dansk Botanisk Arkiv 7: 55, 56, t. 11, figs 10-12 (Oct.1932).— Type: Madagascar, Mandraka, Aug. 1906, d’Alleizette 111 (holo-, BM!); Imerina: Betsitra, près Andsoforobé (sic), Oct. 1906, d’Alleizette 170 (para-, BM).
OTHER MATERIAL EXAMINED. — Réunion. Insul. Borboniae, 1845, Garnier 52 (BM).
Madagascar. Ankafana, 1880, Cowan s.n. (BM000800963, BM000800964,BM000800965).—Mandraka, VIII.1906, d’Alleizette 111 (BM). — Massif du Kalambatitra (Centre- Sud), Mont Analatsitendrika, forêt ombrophile (sur latérite de gneiss), 1650-1800 m, XI.1933, Humbert 11948 (P00349504, P00349505). — Massif du Kalambatitra (Centre-Sud), mont Beanjavidy, forêt ombrophile, alt. 1500-1700 m, XI.1933, Humbert 12067 (P00349506).— Montagnes entre le haut Sambirano et le haut Maivarano (entre Mangindrano et Ampanompia), forêt ombrophile sur latérite de gneiss, 1400-1800 m, XI.1937, Humbert 18143 (P00349507). — Partie occidentale du massif de Marojejy (Nord-est) de la vallée de l’Ambatoharanana au bassin supérieur de l’Antsahaberoka, forêt ombrophile sur argile latéritique de gneiss et granite, c. 1400 m, 9.XI-2. XII.1959, Humbert & Saboureau 31602 (P00349508). — Fianarantsoa,Ranomafana-Ifanadiana, PN de Ranomafana, forêt de Vatoharanana, à 4 km au sud-ouest du village de Ranomafana, 980 m, 3.X.2000, Rakotondrainibe 5820 (P00181240, P00181241, P00181242). — Fianarantsoa, Ivohibe, dans la RNI d’Andringitra, au nord-ouest d’Ambarongy, au sud-ouest du campement no. 3, 1210 m, 28.XI-1.XII.1994, Rakotovao 267 (P00116711).
DESCRIPTION
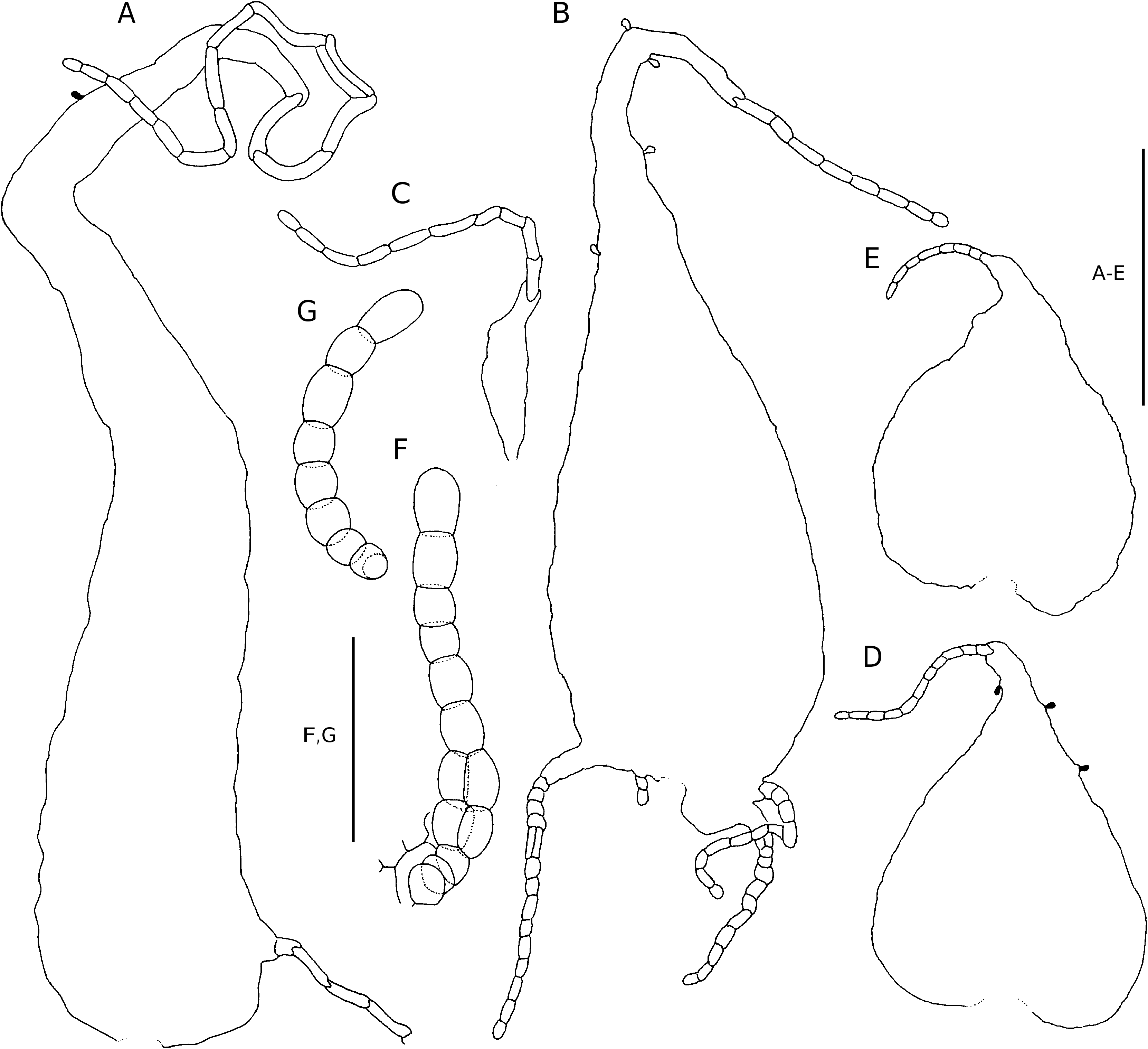
Plants terrestrial. Rhizome short-decumbent or short and erect, up to 15 mm in diameter, set with roots, crowded stipe bases, and scales, the scales ferrugineous, chartaceous, broadly attached, subulate to narrowly lanceolate, up to 30 × 4 mm, entire, or with a few scattered filiform outgrowths near the apex, the apex short-filiform, terminating in an oblong cell. Fronds caespitose, arching, up to 1.5 m long; stipe base castaneous, stramineous higher up, sulcate adaxially, up to 815 mm long and 9 mm in diameter, proximally densely scaled, subglabrous higher up, the scales ferrugineous to castaneous, chartaceous and often rugate, or thinly crustaceous, broadly attached or sessile, filiform to broadly ovate, up to 40 × 9 mm, truncate to cordate, entire, or with a few scattered filiform outgrowths near the base or apex, occasionally with a few scattered glands (these frequently also occur on the scale laminae), and short uniseriate hairs, the scale apex filiform, terminating in a short uniseriate series of cells, the apical cell oblong or spheroidal, higher up often with glands; lamina herbaceous, broadly ovate, to 4-pinnate-pinnatifid, anadromous, catadromous towards the apex, up to 625 mm long, with up to 14 pairs of petiolated pinna pairs; rachis stramineous, adaxially shallowly sulcate, becoming narrowly winged towards the apex, sparsely scaled, the scales castaneous to ferrugineous, chartaceous, to thinly crustaceous, broadly attached or sessile, filiform to broadly ovate, up to 6 × 3 mm, truncate to cordate, the margins regularly set with glands or with a few uniseriate hairs near the base, the apex filiform, terminating in a short uniseriate series of cells, the apical cell oblong or spheroidal; pinnae petiolate, the petiole up to 22 mm long, the basal pair inaequilaterally ovate, ovate to lanceolate towards the lamina apex, to 3-pinnate-pinnatifid, the basal pair the longest, basiscopically developed, up to 345 × 230 mm, opposite to alternate, basally widely spaced, more closely spaced apically, often imbricate, with up to 12 petiolated pinnule pairs; pinna-rachis adaxially shallowly sulcate, the sulcus confluent with that of the rachis, often closely set with unicellular glandular hairs and scales, abaxially sparsely scaled, the scales ferrugineous to stramineous, chartaceous, sessile, ovate to broadly ovate, up to 4.5 × 2.7 mm, cordate, entire or regularly set with glands, the apex filiform; pinnules petiolate, the petiole up to 9 mm long, ovate, to 2-pinnate-pinnatifid, the acroscopic pinnule on the basal pinnae up to 105 × 29 mm, the basiscopic pinnule on the basal pinnae up to 150 × 70 mm, opposite to alternate, spaced or slightly imbricate, with up to 9 petiolated segment pairs; pinnule-rachis adaxially sulcate, sparsely scaled, the scales similar to, but smaller than those on the pinna-rachis; segments petiolate, the petiole up to 1 mm long, ovate, to 1-pinnatepinnatifid, with up to 3 pairs petiolated ultimate segments, spaced, the basiscopic segment on the basal pinnae up to 44 × 20 mm, the acroscopic pinnule on the basal pinnae up to 28 × 14 mm; costa adaxially sulcate, winged, ridged adaxially, abaxially sparsely scaled, the scales similar to, but smaller than those on the pinna-rachis; ultimate segments oblong-obtuse, firmly herbaceous, the basiscopic ultimate segment on the basal segment up to 11 × 6 mm, the acroscopic ultimate segment on the basal segment up to 9 × 6 mm, deeply lobed, the lobes serrate to obtusely dentate, glabrous adaxially, abaxially glabrous or often with glands along the veins, (46-)66(-84) mm long, with isocytic to moniliform hairs and scales, the scales often bullate, sessile, broadly ovate, up to 1.5 × 0.7 mm, truncate to cordate, entire or with a few scattered glands, the apex filiform, terminating in an oblong or spheroidal cell.Venation anadromous, becoming catadromous towards the lamina and pinna apex, pinnately branched in the segments, the vein branches pinnately banched, forked or simple near the apex, evident, ending in the teeth near the margin. Stomata mostly of the polocytic type, (30-)39(-50) mm long. Sori circular, inframedial on the anadromous vein branches, discrete at maturity, up to 1 mm in diameter; sporangium stalk simple, glandular, or rarely with a pluricellular uniseriate hair, capsule with (12-)13(-16) indurated annulus cells, epistomium (4-)5(-6)-celled, hypostomium (4-)6(-7)-celled; indusium brown, firmly herbaceous, reniform, entire or glandular along the margin (rarely also on the laminae), up to 1 mm in diameter.Spores ellipsoidal, monolete, the perispore with low tubercules and/or long reticulate ridges, minutely rugulose to minutely scabrous, (32-)36(- 42) × (20-)22(-26) mm (Tryon & Lugardon 1990: 426, fig. 159.19). Chromosome number: 2n = c. 82 (Vida in Widén et al. 1973: 2129).
REMARK
The correct spelling of the specific epithet is discussed by Pichi Sermolli (1985: 158).
DIAGNOSTIC FEATURES AND RELATIONSHIPS Dryopteris kilemensis can be separated from all other African Dryopteris species by its large structure, fronds up to 1.5 m long, and the finely dissected lamina, which may be up to 4-pinnate-pinnatifid. The sessile, broadly ovate, often bullate scales with the apex terminating in a short uniseriate range of cells are also characteristic.
VARIATION
Dryopteris kilemensis shows little variation in gross morphology, but micromorphological variation was detected in scale morphology, the presence of glands on the lamina, the presence of glands or pluricellular hairs on the sporangium stalk, and the distribution of glands on the indusium lamina.
Scale texture appears to be influenced by environmental conditions, as plants occurring in less favourable conditions have scales that are thinly crustaceous rather than chartaceous.The rhizome and frond scales of these plants are generally also broader than those of plants from moist forest habitats. Glands and pluricellular hairs along the scale margins also show variation in frequency.
A direct correlation cannot be drawn between habit and the presence of glands occurring on the lamina. Whilst some collections from higher elevations bear these structures, others do not. Similarly, glandular collections from moist forests were also observed.
The sporangium stalk of D. kilemensis is generally simple, but in some the sporangia bear a single gland on the stalk.In others,a long simple pluricellular hair is borne near the stalk base.No collections were observed where both glands and pluricellular hairs occur.
Generally, the indusium of D. kilemensis is simple, or it may bear a number of glands along the margin. The occurrence of glands on the indusium lamina is, however, less frequent and have only been observed in a few collections from Africa.
DISTRIBUTION AND HABITAT
Dryopteris kilemensis is confined to the central and eastern parts of tropical Africa and Madagascar
where it occurs in forests at 980 to 1800 m ( Fig. 27 View FIG ). Although reported from Réunion, it has not been collected on that island recently.
SPECIES INCERTAE SEDIS
Fraser-Jenkins (1986: 198) places Dryopteris wardii in subgenus Nephrocystis sect. Purpurascentes , a position I cannot support. The unique features the species exhibits (see Diagnostic features and relationships) cast a doubt over its inclusion in Dryopteris , but due to the lack of a better placement, it is here tentatively retained in this genus. A molecular and detailed morphological studies based on live material are required to ascertain the correct placement of D. wardii .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dryopteris kilemensis (Kuhn) Kuntze
| Roux, Jacobus P. 2011 |
Nephrodium lastii
| Baker 1891: 324 |