
Protothymallus, Laube, 1901
|
publication ID |
https://doi.org/ 10.5252/g2013n1a3 |
|
DOI |
https://doi.org/10.5281/zenodo.4818172 |
|
persistent identifier |
https://treatment.plazi.org/id/817987A0-577C-B003-79A3-B0EC0A36FD83 |
|
treatment provided by |
Felipe |
|
scientific name |
Protothymallus |
| status |
|
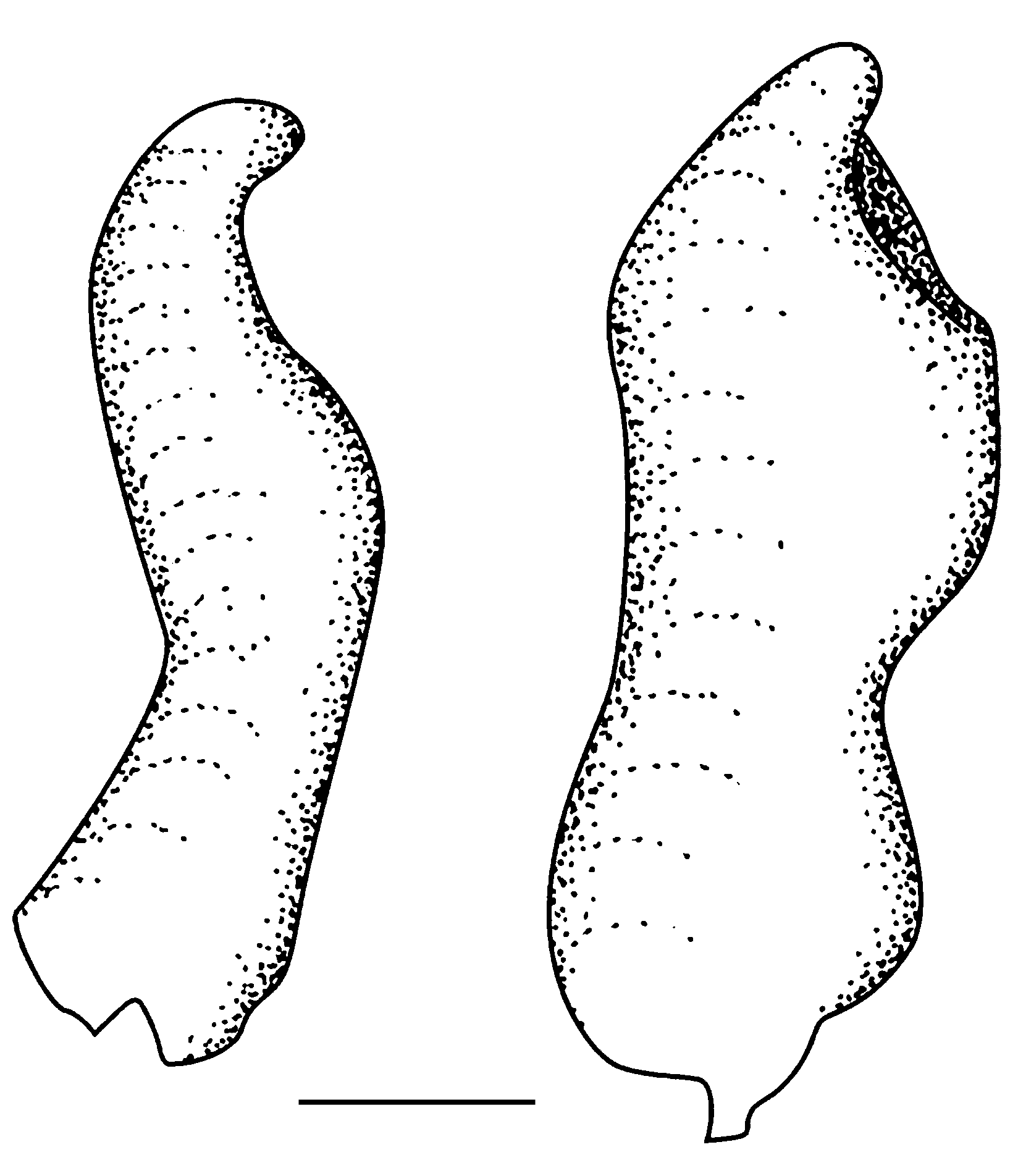
Protothymallus sp. vel Varhostichthys sp. ( Figs 13 View FIG ; 14 View FIG )
DESCRIPTION ANATOMIQUE
Lors des travaux de terrassement réalisés en 1981 pour la construction d’un hypermarché du Cen-
tre commercial ouest, nous avons recueilli deux spécimens incomplets attribuables soit au genre Varhostichthys Obrhelová, 1969, soit à Protothymallus Laube, 1901 . Malheureusement, ces deux spécimens sont dépourvus de tête.
Comme le montre le spécimen le plus complet, le corps est élancé. Sa hauteur maximale devait égaler environ un quart de la longueur standard, laquelle était un peu inférieure à 150 mm. La colonne vertébrale se compose de 40 ou 41 vertèbres dont 24 ou 25 constituent la région abdominale (y compris les centra modifiés pour former l’appareil de Weber). Le nombre de vertèbres postabdominales est de 17 sur l’un des spécimens et seulement de 15 sur l’autre. Les centra vertébraux supportent des neurapophyses et des hémapophyses bien développées dont l’extrémité distale atteint, dans la région abdominale, les ⅔ de la distance séparant les parties dorsale et ventrale des centra, respectivement des bords dorsal et ventral de l’animal. Des epineuralia et des epipleuralia sont observables dans la région postabdominale. Les premiers sont également présents dans la région abdominale. Quelques traces de dorsospinalia sont visibles entre les premières neurapophyses abdominales. On remarque en particulier la présence du premier d’entre eux, en forme de lame aplatie dans le plan sagittal, en avant de la neurapophyse portée par le premier centrum abdominal libre. Les côtes pleurales sont au nombre de 15 ou 16 paires. Elles sont très développées puisque, à l’exception de celles situées en arrière des nageoires pelviennes, leur extrémité distale atteint pratiquement le bord ventral de la cavité abdominale. En avant de la première paire de côtes pleurales s’observe la
Gaudant J.
paire de parapophyses très développées portées par le quatrième centrum modifié pour constituer l’appareil de Weber.
La nageoire caudale est assez grande puisque sa longueur devait être comprise un peu moins de cinq fois dans la longueur standard. Elle est profondément bifurquée, la longueur de ses lépidotriches axiaux n’excédant pas la moitié de celle des plus longs rayons de chaque lobe. Elle se compose de 19 rayons principaux, dont 17 bifurqués, en avant desquels prennent place, dorsalement et ventralement, respectivement 9 et 8 rayons marginaux.
La nageoire caudale est soutenue par un squelette caudal axial composé de quatre vertèbres. On dis - tingue à l’arrière le complexe uro-terminal, prolongé par le pleurostyle qui est redressé d’environ 40° par rapport à l’axe longitudinal de la colonne vertébrale. Le complexe uro-terminal supporte ventralement le parhypural et cinq hypuraux. Il est surmonté par un arc neural réduit auquel fait suite un épural unique. En avant du complexe uro-terminal, trois centra préuraux libres portent des neurapophyses et hémapophyses allongées qui sont également plus ou moins élargies pour soutenir les rayons marginaux dorsaux et ventraux de la nageoire caudale.
La nageoire dorsale était probablement insérée un peu en arrière du milieu du corps et légèrement en arrière de la verticale passant par l’origine des pelviennes. Elle débute par deux rayons courts en arrière desquels prend place un long rayon articulé qui est le plus développé de la nageoire et dont la longueur représente approximativement les ¾ de la hauteur maximale du corps. Plus en arrière, on dénombre sept lépidotriches à la fois articulés et bifurqués dont la longueur diminue progressivement vers l’arrière.
L’endosquelette de la nageoire dorsale se compose de huit ptérygiophores. À l’exception du premier d’entre eux qui est bifide, les ptérygiophores sont constitués d’un épaississement osseux longitudinal sur lequel prend appui une lamelle osseuse triangulaire disposée dans le plan sagittal.
La nageoire anale occupe une position reculée. Composée de neuf rayons, elle débute par deux rayons courts en arrière desquels prend place un long rayon articulé qui n’est pas bifurqué. La partie postérieure de la nageoire est composée de six (ou sept?) lépidotriches à la fois articulés et bifurqués dont la longueur diminue assez rapidement vers l’arrière.
L’endosquelette de la nageoire anale est formé de six ou sept ptérygiophores. Les premiers d’entre eux, très longs, s’intercalent profondément entre les hémapophyses correspondantes.
Les nageoires pectorales, qui sont composées de 17 rayons, sont insérées en position basse sur le flanc. Elles sont de taille modérée puisque l’extrémité distale de leurs plus longs rayons ne dépasse qu’à peine le milieu de l’espace séparant la base des pectorales de celle des pelviennes.
Les nageoires pelviennes, situées un peu plus près de l’origine de l’anale que de la base des pectorales, sont relativement grandes puisque la longueur de leurs rayons les plus développés est approximativement égale à celle des plus longs rayons des pectorales. Elles semblent avoir été constituées de neuf rayons, dont huit à la fois articulés et bifurqués, auxquels s’ajoute dorsalement un très court rayon accolé au rayon non bifurqué.
Les écailles, de type cycloïde, paraissent avoir été relativement grandes. Leur surface porte de nombreux circuli concentriques dont la densité semble avoir été proche de 25 au millimètre. En outre, une douzaine de radii ornent le champ postérieur de l’écaille, alors que le champ antérieur de l’écaille semble en avoir été dépourvu. La ligne latérale est observable dans la région postabdominale. Elle prend place un peu audessus de la colonne vertébrale qu’elle rejoint vers l’avant au niveau des dernières vertèbres abdominales.
Deux dents pharyngiennes isolées du type « en crochet » (« Hakenzahn » de Rutte 1962) ont été recueillies près des débris de la tête du second spécimen ( Fig. 14 View FIG ). L’une est une dent postérieure effilée, de forme sigmoïde. La seconde possède une couronne renflée au-dessous de l’area masticatrice qui détermine un angle de 40° avec l’axe de la couronne.
DISCUSSION
L’ensemble des caractères décrits précédemment permet d’affirmer que ces deux fossiles appartiennent bien à la famille des Cyprinidae comme l’indiquent notamment la disposition relative des nageoires, la structure du squelette caudal axial, la possession d’un appareil de Weber dont témoigne la présence des longues parapophyses en avant de la première paire de côtes pleurales. Au sein de cette famille, la composition de la colonne vertébrale, formée de 24 ou 25 vertèbres abdominales et de 15 à 17 vertèbres postabdominales, conduit à attribuer ces poissons soit au genre Varhostichthys Obrhelová, 1969, soit à Protothymallus Laube, 1901 . On notera en effet que les deux spécimens d’Aix-en-Provence possèdent un corps plus élancé que les représentants de l’espèce type Varhostichthys brevis (Agassiz, 1839), de l’Oligocène inférieur de Valeč ( Bohême). En effet, la hauteur maximale de leur corps égale environ le quart de la longueur standard alors que, chez Varhostichthys brevis (Agassiz, 1839), elle atteindrait 40 % de celle-ci selon Böhme (2007) et seulement un tiers selon Gaudant (2009). Or, la hauteur maximale du corps est généralement comprise entre 20 et 25 % chez Protothymallus elongatus (Kramberger, 1885) , une espèce de Cyprinidae de l’Oligocène de Bohême et de Saxe ( Allemagne) qui possède une colonne vertébrale comportant 23-24 vertèbres abdominales et 15- 16 postabdominales, et des nageoires dorsale et anale composées respectivement de (9) 10-11 rayons et de 9-10 rayons ( Gaudant 2009). Les caractères méristiques de cette espèce sont donc très proches de ceux observés sur les deux spécimens d’Aix-en-Provence. L’absence probable de radii sur le champ antérieur des écailles incite donc à penser que les deux spécimens décrits ci-dessus pourraient être des représentants du genre Protothymallus Laube, 1901 car, comme l’a montré Böhme (2007), l’espèce Varhostichthys brevis (Agassiz, 1839) possède des écailles dont les champs antérieur et postérieur sont tous deux ornés de radii.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Family |
Protothymallus
| Gaudant, Jean 2013 |
Protothymallus
| Laube 1901 |