
Brasilotitan nemophagus, Machado, Elaine B., Avilla, Leonardo Dos S., Nava, William R., Campos, Diogenes De A. & Kellner, Alexander W. A., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3701.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:73622FCA-CCDA-4A7D-A036-530B6B120E1E |
|
DOI |
https://doi.org/10.5281/zenodo.5620345 |
|
persistent identifier |
https://treatment.plazi.org/id/7D17D258-637B-A410-FF0D-C5878CAAD6CD |
|
treatment provided by |
Plazi |
|
scientific name |
Brasilotitan nemophagus |
| status |
sp. nov. |
Brasilotitan nemophagus new species
Etymology. Composed by the union of two words from the Greek: némos, meaning pasture, wood + phagos, meaning to eat, in allusion to the herbivorous nature of this species.
Holotype: Right dentary, two cervical vertebrae, three incomplete sacral vertebrae, fragment of an ilium, fragments of an ischium, one ungual and fragmentary elements, all housed at the Museu de Paleontologia de Marília (MPM 125R). Casts of the dentary are housed at the Museu Nacional/UFRJ (MN 7371-V).
Referred material: an isolated tooth housed at the Museu de Paleontologia de Marília (MPM 126R).
Locality and horizon: The material was collected at the kilometer 571 (22o07’04S, 51o26’33W) of the Raposo Tavares state road (SP-270), near Presidente Prudente city ( Fig 1 View FIGURE 1 ), São Paulo State, Brazil. It was recovered from a reddish fine-grained sandstone that is part of the Adamantina Formation, whose age is regarded as Turonian- Santonian (Dias-Brito et al. 2001) or Campanian-Maastrichtian age (Gobbo-Rodrigues et al. 1999).
Diagnosis. titanosaur sauropod with following autapomorphies: (1) dorsal symphyseal region of the dentary twisted medially; (2) dorsal portion of the symphyseal contact thicker anteroposteriorly than the ventral part; (3) presence of anteriorly directed accessory prezygapophyseal articulation surface on cervical vertebrae; (4) intraprezygapophyseal laminae of cervical vertebrae V shaped in dorsal view.
Brasilotitan nemophagus can further be distinguished from other titanosaurs by the following combination of characters: mandible with an ‘L’ shaped morpology in dorsal view; sharp anteroposteriorly oriented crest on the dorsal margin at the posterior region of the mandibular ramus; cervical vertebrae with dorsoventrally low neural spines (approximately 36% and 38% of the total size of the vertebrae); ventral portion of the cervical centrum arched dorsally; prezygapophyses of cervical vertebrae are placed anteriorly relative the anterior articulation of the centrum.
Description and comparisons. Overall the material of Brasilotitan nemophagus is preserved with little or no distortion, a typical feature of sauropod bones from the fossiliferous layers of the Bauru Basin (e.g. Kellner & Campos 1999). Most elements are broken or incomplete, with the cortical bone well preserved despite being broken in several parts.
Dentary. The surface of the right dentary is weakly ornamented with pits, grooves, and foramina. In dorsal view, this bone is ‘L’ shaped ( Fig. 2 View FIGURE 2 A), with two distinctly oriented portions that meet at an almost right angle. This produces the so-called squared jaw morphology of the lower jaw as seen in Bonitasaura , Antarctosaurus wichmannianus and some diplodocoids (Upchurch 1995), but differing from the more rounded condition reported in Tapuiasaurus , Rapetosaurus , Nemegtosaurus , and Malawisaurus (Wilson 2005; Zaher et al. 2011). Close to the contact surface with the opposite dentary, the mandibular ramus is twisted with the dorsal part directed medially, which has not been reported in any titanosaur before. Right at the contact, this bone reaches its deepest part dorsoventrally ( Table 1 View TABLE 1 ).
A total of 14 alveoli are recognized, more than in Rapetosaurus (11) and Nemengtosaurus (13), but less than in Malawisaurus and Antarctosaurus , where a total of fifteen and sixteen alveoli are reported, respectively (Jacobs et al. 1993; Curry Rogers & Forster 2004; Gallina & Apesteguia 2011). In Bonitasaura , the lower jaw is incomplete lacking the anterior end and therefore the total number of teeth is unknown, while in Tapuiasaurus , despite its completeness, this information was not presented (Zaher et al. 2011).
None of the alveoli in Brasilotitan can be observed in lateral or anterior view since they are hidden by a developed and thin bony wall that rises at the labial margin of the dentary. This structure was termed lateral plate and is observed in all eusauropods (Upchurch 1995). Between the eleventh and fourteenth alveoli the lateral wall of the dentary is crushed and pushed medially. The first five alveoli are the largest and more squared-shape in occlusal view. More posterior alveoli slightly decrease in size.
Dental foramina (= replacement alveoli) are present near the lingual border of each alveolus. Some show the tips of unerupted replacement teeth. The first five dental foramina are very small, while the sixth is larger and circular. Overall, while the alveoli tend do decrease in size posteriorly, the dental foramina tend to increase in size.
Nutrient foramina are also observed, piercing the external surface of the dentary ( Fig. 2 View FIGURE 2 B–C). The first seven anterior foramina are deep, circular, regularly spaced, and are placed close to the ventral border of this bone. Towards the posterior and lateral margin of the dentary, there are three foramina that decreases in size and are less conspicuous, with variable position and spacing.
Posterior to the tooth row, the dorsal border of the dentary forms a thin and laterally compressed crest. Such a structure is also found in Bonitasaura , Rapetosaurus and Antarctosaurus wichmannianus , where it is called the guillotine or posterior crest (Apesteguía 2004, Gallina & Apesteguía 2011). The medial surface of the crest is rugose, contrasting with the finely striated lateral surface. At the base on the lateral surface of this crest there is a developed foramen, from which departs a faint, posteriorly directed groove.
A well-developed groove on the ventral surface of the mandible originates at the symphyseal region and extends towards the posterior region reaching the area corresponding to the ninth alveolus. At the posterior end of the mandibular rami, it is possible to observe the Meckelian groove (or channel). Although the external part of the dentary is broken, the internal wall of the Meckelian channel is complete, indicating that it is open medially. A splint-like broken bone present inside this channel might be part of the splenial. There is a knob-like thickened area in this bone that might be pathological ( Fig. 2 View FIGURE 2 D).
The contact surface for the opposite dentary is P-shaped, with the labial margin straight and the lingual margin showing an anteroposteriorly expanded dorsal and a slender ventral portion ( Fig. 2 View FIGURE 2 E). The dorsal expansion is quite rugose contrasting with the smooth ventral part. Since there is no evidence of breakage in this region, the right dentary was not fused with the left one (not found) to form a mandibular symphysis.
Teeth. No complete tooth is present in the dentary, but fragments can be recognized in some alveoli or in the dental foramina immediately below, what is the case of the fourth, fifth, ninth, twelfth; and fourteenth tooth position. The most complete is one portion of an emerging tooth found in the fourth alveolus that shows a subcircular transverse section but clearly indicates the presence of lateral carinae ( Fig. 2 View FIGURE 2 F). CT Scan images show the presence of at least two other teeth in the fourth alveolus ( Fig. 3 View FIGURE 3 ), totaling three, with the first one being a functional tooth, and the other replacement teeth. CT Scan images also show that in the seventh alveolus, there are two replacement teeth (the functional tooth might have fallen out before final burial and fossilization).
Due to the particular shape of the dentary, the first seven teeth must have faced anteriorly while the seven remaining ones would have faced laterally (including the eighth that had an intermediate position). In the two other titanosaurs with similar square-shaped lower jaw, Bonitasaura and Antarctosaurus wichmannianus , the number of laterally facing teeth are five and seven, respectively.
The middle portion of an isolated tooth (MPM 126R) was found associated with the type material of Brasilotitan nemophagus and might belong to this species or perhaps even to same specimen ( Fig. 4 View FIGURE 4 ). This tooth has a circular transverse section, with finely striated crown. There is also the indication of carinae, which are not symmetrical, somewhat displaced lingually, suggesting that this tooth might have been located at the region where the two distinctly oriented portion of the dentary met. Isolated sauropod teeth, including those attributed to titanosauriforms, are common and reported in several deposits, (e.g. Saegusa & Tomida 2011) having overall a similar morphology.
Cervical vertebrae. Only two incomplete cervical elements from Brasilotitan nemophagus were recovered ( Figs 5–9 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ). In both the neural spines is single (e.g. not bifid), transversally thickened and a blunt tip, bordered by a deep lateral depression. Both vertebra possess camellate internal structure.
The smallest cervical vertebra lacks the anterior part and, based on the elongated and slender centrum and the dorsoventrally short neural spine, likely belongs to the anterior portion of the neck. The centrum is extremely arched dorsally, much more than in the anterior cervicals of Pitekunsaurus macayai Filippi & Garrido, 2008, with the ventral margin appearing strongly concave in lateral view (Filippe & Carrido 2008). The ventral surface tends to be flattened with the lateral surfaces straight, making the outline of the centrum rectangular, differing from the more cylindrical condition observed in some other titanosaurs (e.g. Trigonosaurus ). It lacks the marked depression seen in the ventral margin near the parapophyses of Uberabatitan, Trigonosaurus and MCT 1487-R (Salgado & Carvalho 2008; Campos et al. 2005). A small depression bounded by anteroposteriorly oriented ridges is present on the right lateral surface dorsal to the parapophysis. On the left, this depression is less conspicuous and divided by irregular oriented bony struts. Two longitudinal ridges are present on the ventral surface, which are well-marked, particularly near the cotyle.
The cotyle is concave with the ventral border displaced posteriorly relative to the dorsal one. The posterior exit of the neural channel is broad and reniform, with the horizontal axis longer than the vertical one. There is a constriction of the base of the neural spine above the neural channel. The zygapophyses are not preserved.
In both sides the cervical rib loop (ansae costotransversariae; Wedel & Sanders, 2002) is broken since the transverse processes, the cervical ribs and the parapophyses are not complete. The postzygodiapophyseal lamina (PODL) is well developed, contrasting with the much thinner posterior centrodiapophyseal lamina (PCDL). The region where those two laminae meet houses the postzygapophyseal centrodiapophyseal fossa (pocdf).
The spinopostzygapophyseal lamina (SPOL) is approximately as robust as the spinoprezygapophyseal lamina (SPRL), which is not complete in this cervical vertebra. Due to the small extension of the neural spine, the SPOL and the SPRL are essentially continuous. This condition differs from the cervical vertebrae of Trigonosaurus, Maxakalisaurus and Uberabatitan (Campos et al. 2005; Kellner et al. 2006; Salgado & Carvalho 2008), in which those laminae are not confluent. The postzygapophyseal spinodiapophyseal fossa (posdf) is very well-developed.
The second cervical vertebra presents the anterior half, including parts of the cervical ribs. It is larger, more robust and has a dorsoventrally higher neural spine indicating that it represents a more posterior element in the neck compared with the previous one. However, compared with other titanosaurids the neural spine of Brasilotitan is shorter (e.g. Kellner et al. 2006; Calvo et al. 2007; Salgado & Carvalho 2008; Table 2 View TABLE 2 ).
The condyle is partially preserved and, despite being laterally compressed, clearly was highly convex. The centrum is incomplete and some parts have been distorted. Nevertheless, it shows a well-developed ventral depression between the parapophyses, as in Trigonosaurus, Uberabatitan and MCT 1487-R. On the left side, the lateral surface of the centrum is rather smooth while on the right side the surface is rugose, showing depression and ridges interpreted as muscle scars. The parapophyses are elongated, forming a rather thin blade that apparently extends for over half the centrum length. D'Emic (2012) suggested that the presence of this feature in posterior cervical vertebrae was diagnostic for the titanosaurian clade Lithostrotia. Despite the ventrolateral position of the parapophysis, it does not reach the condition reported for in some euhelopodid taxa (see D’Emic 2012), where it projects ventrally, exceeding the ventral margin of the centrum (e.g. Ksepka & Norell, 2006).
The prezygapophyses are large with flat and slightly dorsomedially directed articular surfaces. They are very well developed and extend beyond the anterior border of the condyle, differing in this respect from the saltasaurines (Powell 1992) and MCT 1487-R. They are also proportionally longer than in Trigonosaurus, Maxakalisaurus, Uberabatitan , Futalognkosaurus and Rapetosaurus . They further differ from MCT 1487-R by being straighter and not curved dorsally.
Ventral to the prezygapophyses, the prezygodiapophyseal lamina (PRDL) is rather pronounced, more than in most titanosaurs (e.g. MCT 1487-R, Trigonosaurus ). This vertebra of Brasilotitan presents a pre-epipophyses, as seen in other sauropods (Wilson & Upchurch 2009). However, above the pre-epipophysis, there is a flat surface, forming an anterior accessory prezygapophyseal articulation, that to our knowledge has not been reported in Titanosauria before.
The SPRL are slightly asymmetrical and do not reach the prezygapophyses or the anterior margin of the intraprezygapophyseal lamina (TPRL). On the dorsal surface of the TPRL, that forms the anterior dorsal margin of the neural canal between the SPRL, a faint groove bordered by delicate ridges is observed.
The epipophyseal-prezygapophyseal lamina (EPRL) originates close to the prezygapophysis, and reaches the base of the neural spine. On the right side, this lamina runs towards the dorsal part of the neural spine, dividing the spinodiapophyseal fossa (sdf), similar to the condition observed in Erketu (Wilson et al. 2011). Oddly, on the left side, the EPRL stops on the basis of the neural spine and does not divide the spinodiapophyseal fossa. In any case the anterior part of the EPRL differs from MCT 1484-R by being blunter.
The intraprezygapophyseal lamina (TPRL), that forms the dorsal border for the neural canal, is welldeveloped, wide and ‘V’ shaped (angle between them: ~75°) in dorsal view. This is quite different from the condition reported in other titanosaurs (e.g. MCT 1487-R, Trigonosaurus, Uberabatitan , Futalognkosaurus , Malawisaurus ). A ‘V’ shaped TPRL is also observed in the diplodocid Kaatedocus from the Morrison Formation (Jurassic, United States of America, Tschopp & Mateus 2012), but differs from Brasilotitan by having a more acute angle (~45°).
The CPRL is a robust lamina. On the medial side, bordering the lateral margin of the neural channel, there are additional laminae and small pneumatic cavities.
Besides the differences presented above, the cervical vertebrae of Brasilotitan can be distinguished from Futalognkosaurus , Rapetosaurus , Mendozasaurus , and in Malawisaurus (specimen Mal-245) by having lower neural arches (Gomani 2005; Calvo et al. 2007; González-Riga et al. 2009). Those four taxa further have a laterally expanded neural spine, not present in Brasilotitan .
Cervical ribs. Some fragmentary cervical ribs have been found isolated or connected with both cervical vertebrae. The best preserved belongs to the left side of the larger cervical vertebra. The shaft of the tuberculum is narrow, while the capitulum is broad. The latter is comparatively thicker, particularly at the anterior margin. Compared to some other titanosaurs (e.g. MCT 1487-R), the tuberculum is more inclined posteriorly, and displaced relative the prezygapophysis and the anterior margin of the neural spine.
The caput of the cervical rib is dorsoventrally expanded and mediolaterally compressed, while the reverse is true for the preserved portion of the posteriorly directed shaft. Pneumatic cavities bordered by thick laminae are present anteriorly and posteriorly. Although incomplete, it is likely that the cervical ribs extend beyond the posterior margin of the centrum.
Sacrum. Three sacral vertebra are preserved, lacking most of the centrum and the tips of the neural spines ( Fig. 10 View FIGURE 10 A). Only the neural arch is preserved, lacking the dorsal border of the neural spines and part of the conjoined laminae. The exact number of sacral vertebrae in this species is unknown. All three are tightly connected, but visible sutures between them indicate that they are not completely fused. The sacral vertebra possesses a camellate internal structure, seen in the broken tips of the neural spines.
The first preserved sacral vertebra is the most complete one. The fossa located posterior to the diapophysis and bounded by the postzygapophysis and the neural spine (posdf) is very large and deep. The neural spine of this vertebrae is fused with the neural spine of the subsequent vertebrae at the base (interneural junction), but dorsally they are largely separated. At the separation point, the cortical bone is very thickened mediolaterally. Several areas on the anterior surface of the SPDL bear small pits and bumps, which also extends to the transverse process, particularly on the right side. The prezygapophysis is short, with an expanded, oval articulation surface that is inclined inwards. As usual in the first sacral elements, the transverse process is more prominent and robust than in more posterior elements. A thin lamina at the dorsal surface of this process connects it to the neural spine, and forms a deep medially located fossa.
The second preserved sacral vertebra possesses well-developed SPRL and SPDL, bordering the prezygapophyseal paradiapophyseal fossa (prpadf), prezygapophyseal spinodiapophyseal fossa (prsdf), and postzygapophyseal spinodiapophyseal fossa (posdf). Those fossae are located above the diapophysis and ventral to the neural spine. The transverse process is less robust and has a wider angle relative to the centrum and is therefore oriented closer to the horizontal plane when compared to the first preserved sacral vertebra.
The last preserved sacral vertebra is the most incomplete of the three, having only its mid-posterior portion preserved but, overall, has the same shape as the previous sacral vertebra ( Fig. 10 View FIGURE 10 B). Some broken portion of the transverse processes of other sacral vertebrae are preserved too, but do not show any particular distinguishing features.
Pelvic girdle. Only a small portion of the right ischium is preserved, corresponding to the middle portion of the shaft ( Fig 11 View FIGURE 11 ). The bone lacks the proximal portion, including the ischium peduncle, and the distal portion. The preserved posterior margin has a rounded edge and is comparatively thick mediolaterally. The lateral surface is concave, while the medial surface is convex.
One small fragment of a thick but laminar bone was found, possible pertaining to the ilium. This fragment shows several puncture marks on its surface, some deep and large while others are shallow and small ( Fig 12 View FIGURE 12. A ). They display a conical shape, getting narrow deeper in the bone. We attribute these marks as probable bite marks made by predators or scavengers.
Ungual phalanx. One ungual phalanx has been found ( Fig 13 View FIGURE 13 A). It is incomplete, with rugose margins perforated by foraminen ( Fig 13 View FIGURE 13 B). It is curved, laterally compressed and slightly asymmetrical, with one side less convex than the other. The less convex side bears a marked sulcus. A tuberosity can be observed on the ventral view, and is displaced to the more convex lateral side ( Fig 13 View FIGURE 13 C). This tuberosity resembles a callus, also seen in others titanosauriforms (see Canudo et al. 2008)
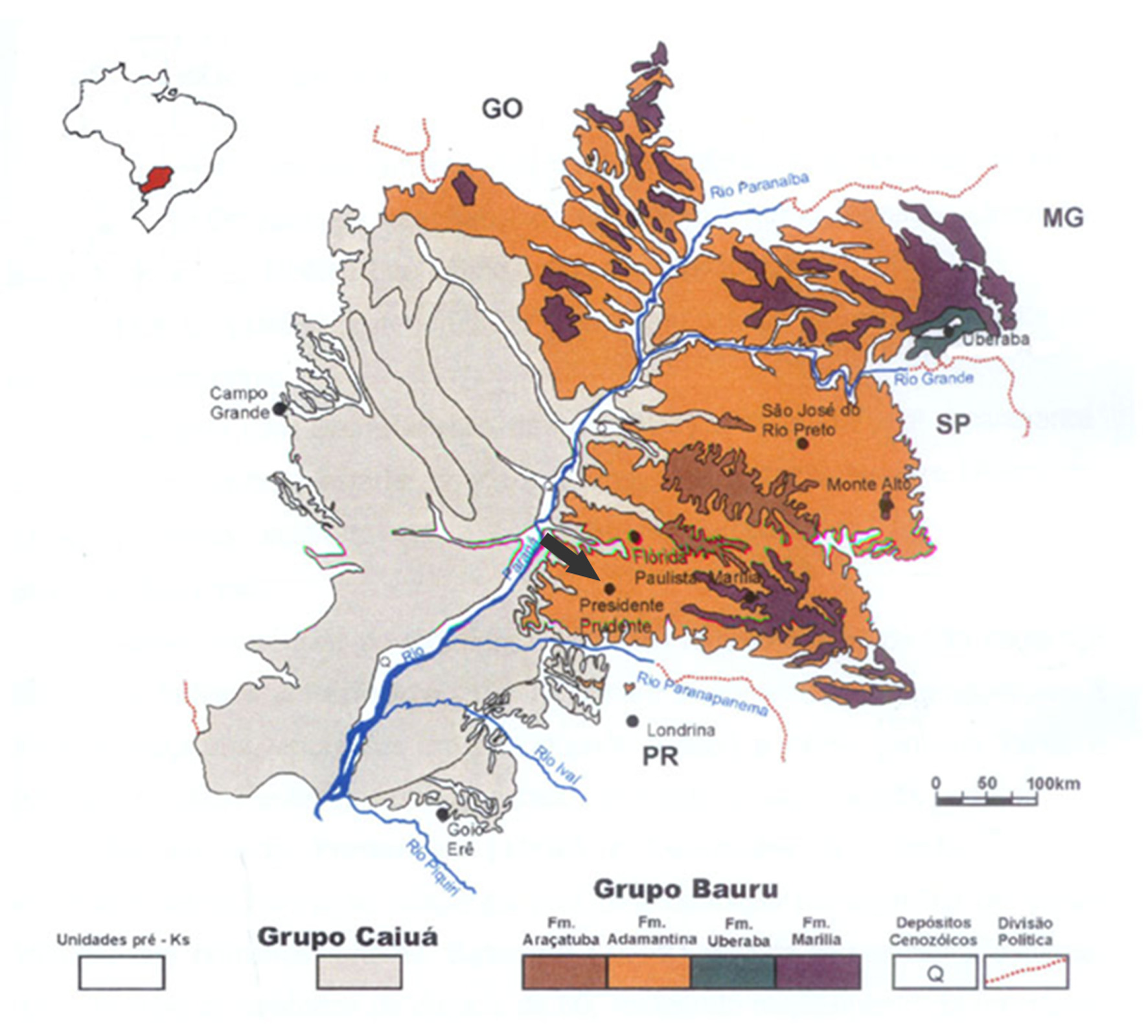





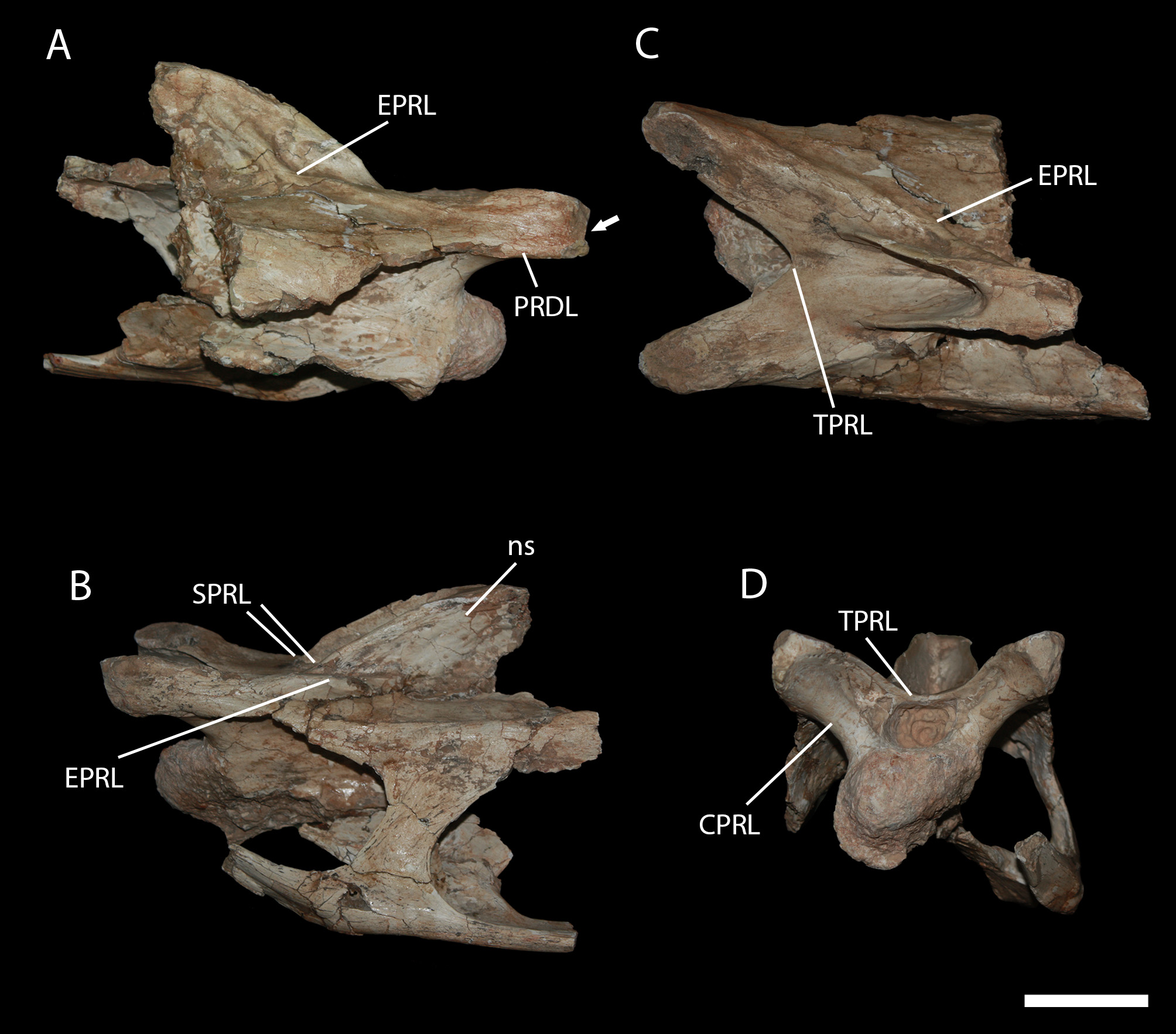


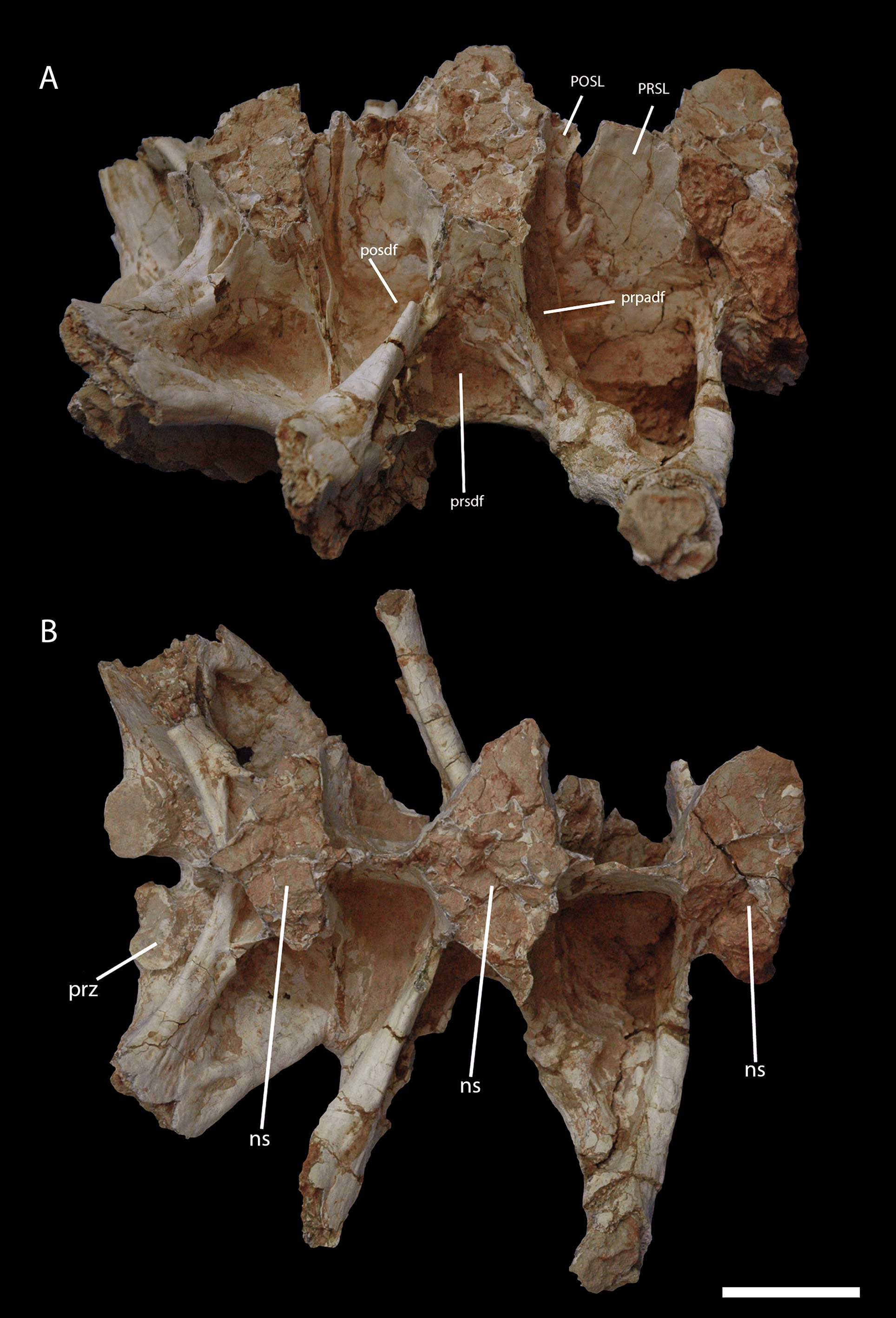



TABLE 1. Measurements in millimeters of the preserved elements of Brasilotitan nemophagus. All measurements were based only on the preserved portions, asterisk indicates incomplete. Centrum and vertebral widths and heights were measured based on the most preserved portions as anteriormost portion for the smallest cervical, and posteriormost for the biggest cervical; for the sacrals, measurements were taken near the neural spine.
| Anteroposterior | Mediolateral | Dorsoventral | |
|---|---|---|---|
| Dentary | 98* (main ramus); 77 (symphyseal ramus) | 21 (at the 9th alveolus) | 38 (at distal end); 34 (at mid length) |
| Isolated tooth | 6 | 5 | 19* |
| Smallest cervical—centrum | 165* | 52 | 38 |
| Smallest cervical—vertebral | 186* | 156* | 117 |
| Biggest cervical—centrum | 147* | 48 | 56 |
| Biggest Cervical—vertebral | 222* | 123* | 153 |
| 1st sacral—neural arch 2nd sacral—neural arch | 102 92 | 201* 272* | 144* 165* |
| 3rd sacral—neural arch | 94* | 21* | 147* |
TABLE 2. Comparative height of cervical neural spines in some titanosaurids.
| total height of the vertebra (mm) | height of the neural spine (mm) | Ratio of heigh of neural spine to total height (%) | |
|---|---|---|---|
| Brasilotitan nemophagus— anterior cervical | 117 | 45 | 38 |
| Brasilotitan nemophagus – midcervical | 153 | 56 | 36 |
| Futalognkosaurus dukei —anterior cervical | 290 | 142 | 49 |
| Futalognkosaurus dukei —midcervical | 953 | 600 | 63 |
| Maxakalisaurus topai— midcervical | 277 | 166 | 60 |
| Maxakalisaurus topai— midposterior cervical | 359 | 192 | 53 |
| Uberabatitan ribeiroi (CPP-1057-UrHo)—anterior cervical | 220 | 85 | 39 |
| Uberabatitan ribeiroi (CPP-1023-UrHo)—midcervical | 280 | 132 | 47 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.