
Paraharmotrema karinganiense, Dutton & DuPreez & Urabe & Bullard, 2022
|
publication ID |
https://doi.org/10.1016/j.ijppaw.2021.09.006 |
|
DOI |
https://doi.org/10.5281/zenodo.10914353 |
|
persistent identifier |
https://treatment.plazi.org/id/7C570C10-3414-8837-FC84-866B7201FB7B |
|
treatment provided by |
Felipe |
|
scientific name |
Paraharmotrema karinganiense |
| status |
sp. nov. |
3.2. Paraharmotrema karinganiense View in CoL n. sp. Dutton & Bullard
3.2.1. Diagnosis of adult specimens (based on six whole-mounted specimens and one hologenophore; USNM coll. nos. 1659278c\, 1659286)
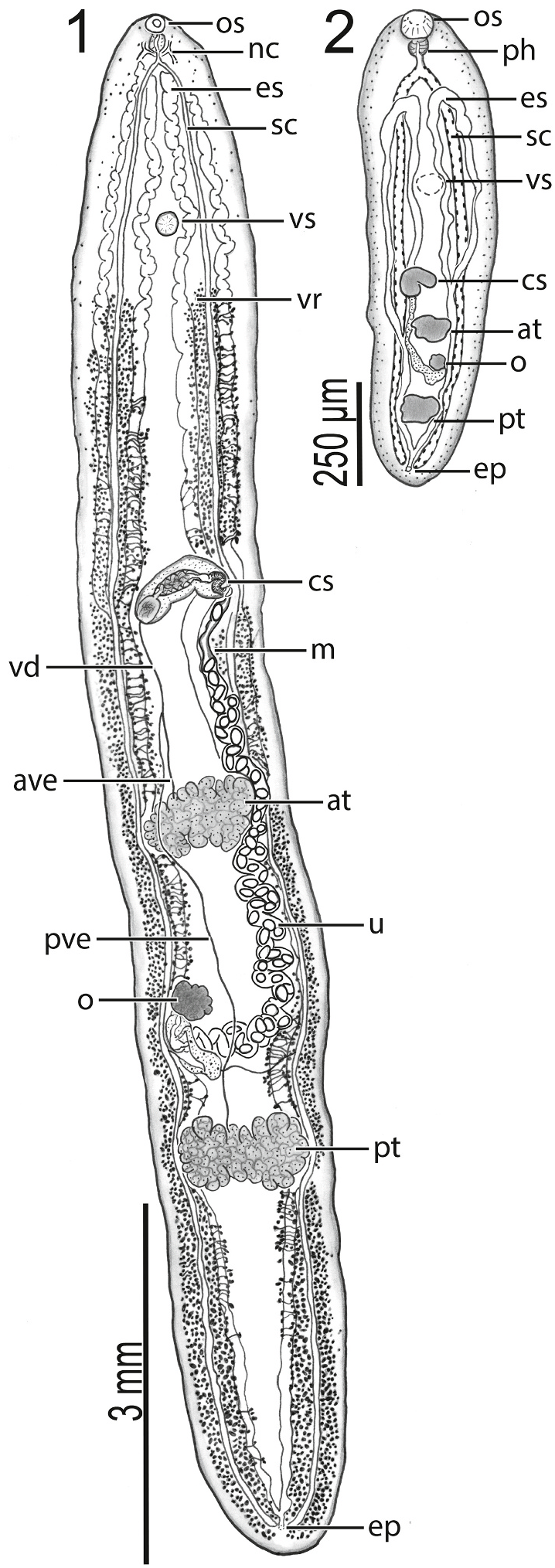
Body 8950 12,675 (10,583 ± 1590; 6) long, 1375 1625 (1504 ± 106; 7) in maximum width at level of ovary ( Fig. 1 2 View Figs ), 5.6–8.5 × (7.1 ± 1.2; 6) longer than wide. Body with ventral concavity in specimens with edges curved ventrally ( Fig. 1 View Figs ). Tegumental scales distributing around anterior end and oral sucker only ( Fig. 7 9 View Figs ). Oral sucker 130–190 (157 ±20; 6) in diameter or 1–2% (1% ± 0%; 6) of body length or 76–94% (87% ±9%; 6) of ventral sucker diameter, 160–180 (168 ±10; 6) wide or 11–13% (11% ±1%; 6) of maximum body width ( Fig. 1 2 View Figs ). Ventral sucker 160–250 (182 ± 34; 6) long or 1–2% (2% ± 0%; 6) of body length, 190–230 (208 ± 16; 6) wide or 28–31% (29% ± 1%; 6) of maximum body width, positioning 1250 1675 (1417 ± 158; 6) or 12–15% (13% ± 1%; 6) of body length from anterior end of body ( Fig. 5 6 View Figs ). Nerve commissure 260–330 (286 ± 27; 6) or 2–3% (3% ± 0%; 6) of body length from anterior body end ( Fig. 1 View Figs ). Pharynx 120–140 (127 ± 8; 6) or 68–80% (71% ±5%; 6) of oesophagus length, 105–135 (118 ± 11; 6) wide or 2–12 × (6 ± 3; 6) wider than maximum oesophagus width ( Fig. 1 2 View Figs ). Oesophagus 150–200 (179 ±17; 6) long, 10–45 (23 ± 12; 6) wide; surrounded by glandular cells. Intestine bifurcating 300–380 (326 ± 30; 6) or 3% (3% ±0%; 6) of body length from anterior body end ( Fig. 1 2 View Figs ).
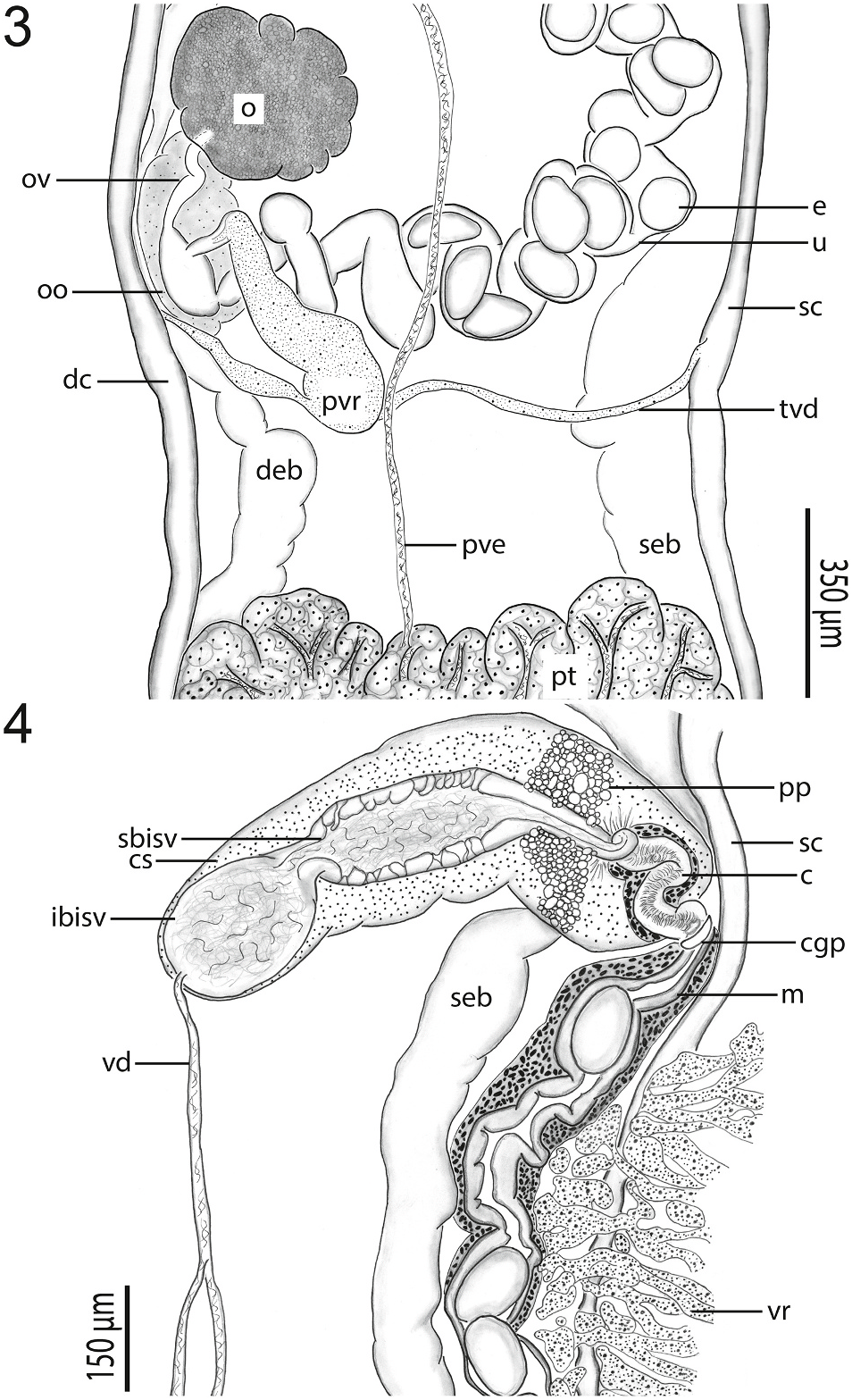
Anterior testis 340 640 (439 ±97; 7) long or 4–5% (4% ±1%; 6) of body length, 680 1000 (797 ±113; 7) wide or 48–62% (53% ±5%; 7) of body width at level of ovary; inter-testicular space 1500 2500 (1889 ± 372; 7) long or 17–22% (18% ±2%; 6) of body length ( Fig. 1 View Figs ). Posterior testis 365 1225 (554 ± 300; 7) long or 3–10% (5% ± 2%; 6) of body length, 640–840 (773 ±68; 7) wide or 39–57% (52% ±6%; 7) of body width at level of ovary, 1925 2950 (2368 ± 393; 7) or 22–24% (23% ±1%; 6) of body length from posterior body end ( Fig. 1 View Figs ). Anterior trunk of vasa efferentia emanating from ventral surface of anterior testis, extending anteriad 400 1150 (724 ±267; 7), or 3–8% (6% ±2%; 6), of body length, 13–25 (21 ± 4; 7) wide; posterior trunk of vasa efferentia emanating from ventral surface of posterior testis, extending anteriad 2000–3780 (2730 ±585; 7), or 22–30% (27% ±3%; 6), of body length, 15–35 (21 ±7; 7) wide, meeting anterior trunk posterior to genital pore between anterior testis and cirrus sac to form vas deferens; vas deferens extending anteriad 350 500 (400 ±58; 7) of body length, 15–25 (20 ± 3; 7) wide before turning sinistrad to insert into the bipartite seminal vesicle ( Fig. 4 View Figs ). Cirrus sac semilunar, 575–900 (671 ± 110; 7) long or 5–7% (6% ± 1%; 6) of body length, 185–300 (216 ± 39; 7) wide or 13–20% (15% ± 2%; 7) body width at level of genital pore; cirrus 188–250 (220 ± 25; 7) long or 25–43% (33% ± 6%; 7) of cirrus sac length, 25–50 (41 ± 10; 7) wide; cirrus spines curved, tapering, 13–25 (20 ± 5; 7) long; cirrus associated with elongate gland cells and numerous prostatic cells ( Fig. 4 View Figs ). Bipartite internal seminal vesicle enclosed within cirrus. Initial bipartite internal seminal vesicle oval, 125–275 (195 ± 52; 7) long or 1–2% (2% ± 0%; 6) of body length, 95–200 (140 ±43; 7) wide, 1–2 × (1 ±0; 7) longer than wide; elongate distal secondary bipartite internal seminal vesicle 205–520 (369 ±100;
46
7) long or 2–4% (3% ± 1%; 6) of body length, 100–140 (109 ± 15; 7) wide, 2–4 × (3 ± 1; 7) longer than wide.
Ovary slightly lobed, 220–350 (266 ±50; 7) long or 2–3% (2% ±0%; 6) of body length, 225–325 (256 ±32; 7) wide or 14–20% (17% ± 2%; 7) of body width; post-ovarian space 2175 4375 (3299 ± 708; 7) or 24–35% (31% ±5%; 6) of body length ( Fig. 3 View Figs ). Oviduct 125–230 (169 ± 37; 6) long or 1–2% (2% ± 1%; 5) of body length, 15–35 (27 ± 8; 6) wide, anterior to transverse vitelline duct, laterally expanding to form ootype ¨. Laurer’ s canal not observed in wholemounts (using SEM, we did observe a pore on the ventral body surface between the level of the ovary and posterior testis that we suspect may be a Laurer’ s canal pore; a 2 μm wide pore accompanied by a tegumental depression having distinctive interconnected ridges). Vitellarium comprising a series of interconnected, irregularly-shaped masses of branched follicles wrapping around caeca and excretory system from posterior to ventral sucker to excretory vesicle, 88–125 (110 ± 15; 7) or 1% (1% ± 0%; 6) from posterior body end; transverse vitelline duct 750 1150 (941 ± 159; 6) in breadth, 15–50 (24 ± 13; 6) wide; primary vitelline collecting duct 325–500 (440 ±58; 7) long, 100–155 (135 ±20; 7) wide, inserting into ootype ¨ventrally ( Fig. 3 View Figs ). O¨otype 100–175 (141 ± 26; 7) long, 15–35 (27 ± 8; 6) wide, between ovary and posterior testis ( Fig. 3 View Figs ). Uterus convoluted, 2600 3970 (3099 ±439; 7) in total length, 100–140 (114 ± 13; 7) wide, extending anteriad around sinistral margin of anterior testis. Eggs longer than wide, 130–160 (148 ±9; 7) long, 85–100 (94 ± 6; 7) wide; number of eggs per specimen 28–55 (41 ±12; 7). Metraterm
47
450–630 (539 ± 75; 7) long, 65–125 (84 ± 19; 7) wide; thick-walled, convoluted, surrounded by small glandular cells ( Fig. 4 View Figs ). Uterus plus metraterm length 3100 4600 (3618 ± 508; 7) or 31–37% (34% ± 2%; 6) of body length. Common genital pore 4550 7900 (6307 ± 1123; 7) or 51–62% (59% ± 4%; 6) of body length from posterior body end, 35–85 (61 ± 16; 7) in diameter, ventral to sinistral caeca ( Fig. 4 View Figs ). Sinistral portion of excretory system 8450 11,135 (9563 ±1165; 6) long, terminating 4100 7600 (5642 ±1210; 6) or 46–65% (53% ±8%; 6) of body length from anterior end; dextral portion of excretory system 8470 11,075 (9683 ± 1161; 6) long, terminating 3000–7435 (4805 ± 1639; 5) or 34–64% (46% ± 13%; 5) of body length from anterior end ( Fig. 1 2 View Figs ).
3.2.2. Taxonomic summary
Type host: Pelusios sinuatus (Smith, 1838) ( Pleurodira : Pelomedusidae ), serrated hinged terrapin; other hosts: Pelusios subniger (Bonnaterre, 1789) ( Pleurodira : Pelomedusidae ), east African black mud turtle; Pelomedusa galeata (Schoepff, 1792) ( Pleurodira : Pelomedusidae ), South African helmeted terrapin.
Site in hosts: Intestinal lumen.
Type locality: Karingani Game Reserve ( KGR) (24 ◦ 20 ′ 8.09 ′′ N 32 ◦ 15 ′ 42.0 ′′ W), Maputo province, Mozambique; GoogleMaps other locality: Roadside borrow pit filled with water (27 ◦ 00 ′ 52.8 ′′ S 32 ◦ 08 ′ 30.1 ′′ E), north-western Zululand, Kwa-ZuluNatal Province, South Africa ( SA) GoogleMaps .
Prevalence and intensity of infections: 2 of 8 (25%) P. subniger from KGR were infected with 22 specimens of the new species; 1 of 3 (33%) P. sinuatus from KGR was infected with 1 specimen ; 4 of 6 (66%) P. galeata from SA were infected with 13 specimens .
Specimens and sequences deposited: Holotype ( USNM 1659278); paratypes ( USNM 1659279 1659286); nucleotide sequences: ex. P. galeata — GenBank No. OL413006, OL413007 ( ITS 2); OL413003, OL413004 (28S); ex. P. subniger — GenBank No. OL413008 OL413008 ( ITS 2); OL413005 (28S); Harmotrema laticaudae ex. L. semifasciata — GenBank No. OL413009 (28S).
Etymology: The specific epithet “ karinganiense ” (neuter) is for the type locality and honors the personnel of KGR for their generous logistic support and cooperation in conducting parasite biodiversity research in Mozambique.
3.2.3. Taxonomic remarks
The new genus can be easily differentiated from the other accepted genera of Liolopidae by the combination of having a linguliform body approximately 6–9 × longer than wide, tegumental spines/scales, a minute ventral sucker located in the anterior 1/7 1/8 of the body,
48
deeply lobed testes that are transverse and abut the caeca (spanning the intercaecal space), a lobed ovary that is dextral and nearest the posterior testis, a uterus that is lateral to the anterior testis (not ventral to anterior testis), and a vitellarium that does not extend anteriad to the level of the ventral sucker and that does not fill the intercaecal space. The new species differs from Helicotrema spp. by having testes in the posterior half of the body; from Liolope spp. by having a cirrus sac that does not abut the ventral sucker; from Harmotrema spp. by having transverse, deeply lobed testes that abut the caeca; and from Dracovermis spp. by having testes that are far apart and that are not limited to the posterior 1/3 of the body ( Table 2 View Table 2 ).
The new species is most similar to species of Harmotrema but further differs from all but one of them by having tegumental spines. The type species of Harmotrema ( H. infecundum ) and all congeners except Harmotrema indica Chattaparhyaya (1970) lack tegumental spines ( Nicoll, 1914; Chattaparhyaya, 1970). Chattaparhyaya (1970) did not measure or draw a spine so we cannot know if the spines are similar/homologous to those of the new genus; we are skeptical that they are present in H. indica . The new species differs from H. indica and all congeners except H. infecundum by having a uterus that is lateral to the anterior testis; H. infecundum has a uterus that was illustrated as lateral to the anterior testis ( Nicoll, 1914). The distribution of vitelline follicles further differentiates these taxa. In the new genus, the vitelline follicles do not extend anteriad beyond nor to the level of the ventral sucker, and the inter-caecal space posterior to the ventral sucker is predominantly void of vitelline follicles ( Fig. 1 View Figs ). Harmotrema infecundum has vitelline follicles anterior to the ventral sucker ( Nicoll, 1914); H. laticaudae has follicles that terminate at level of the ventral sucker and fill the inter-caecal space between the cirrus sac and ventral sucker ( Yamaguti 1933); H. eugari has vitelline follicles that extend to nearly the level of the ventral sucker and fill the inter-caecal space ( Tubangui and Masilungan, 1936); and H. indica has vitelline follicles that fill the intercaecal space between the ventral sucker and cirrus sac as well as between the posterior testis and excretory pore (Chattoparhyaya, 1970). The new genus further differs from Harmotrema spp. by having a large body> 8 mm in length (vs. <7 mm), a minute ventral sucker not spanning the inter-caecal space (vs. ventral sucker spanning inter-caecal space), and a lobed ovary not abutting the posterior testis (vs. ovary not lobed, near to or abutting posterior testis). The new species also has a large uterus that can have> 50 eggs (vs. 21 or less in Harmotrema spp. ).


| USNM |
Smithsonian Institution, National Museum of Natural History |
| SA |
Museum national d'Histoire Naturelle, Laboratiore de Paleontologie |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Digenea |
|
Order |
|
|
Family |
|
|
Genus |