
Cixiinae Spinola, 1839
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4748.1.12 |
|
DOI |
https://doi.org/10.5281/zenodo.3704276 |
|
persistent identifier |
https://treatment.plazi.org/id/7B3B87B8-3602-EA32-FF36-14B90FFD019E |
|
treatment provided by |
Plazi |
|
scientific name |
Cixiinae Spinola, 1839 |
| status |
|
Subfamily Cixiinae Spinola, 1839
Tribe Eucarpiini Emeljanov, 2002
Genus Uphodato Szwedo, 2019
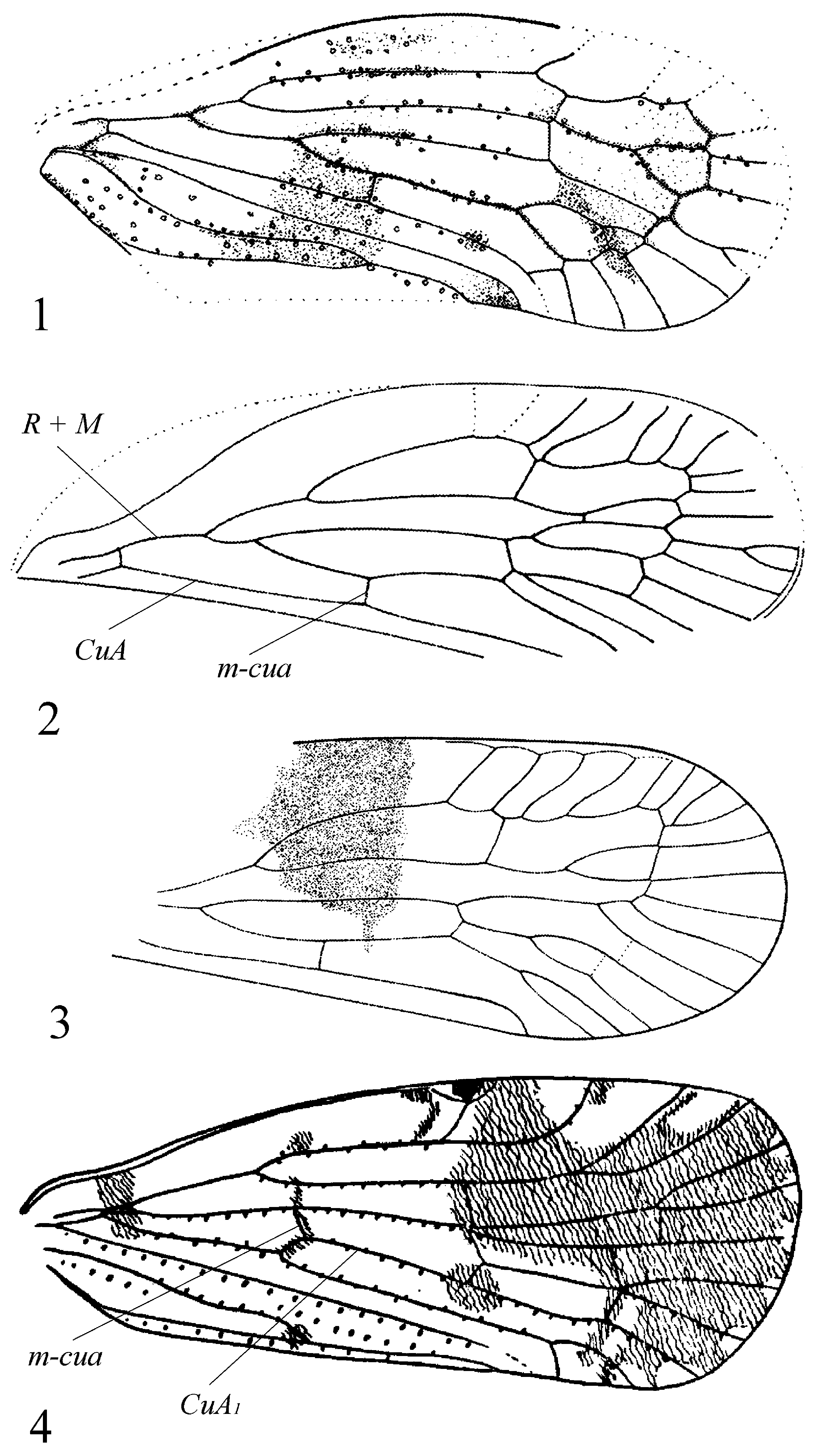
Note. We suggest placement of Breukoscelis Szwedo, 2019 in synonymy under Uphodato Szwedo, 2019 as indicated by the basally concaved costal margin of forewing and peculiar venation of the wing discussed below.
A basal concavity in the costal margin of the forewing is a characteristic feature of the tribe Eucarpiini Emeljanov of Cixiidae and some Achilidae (the genera Tropiphlepsia Muir, 1924 , Apateson Fowler, 1900 , and Achilixius Muir, 1923 ), with their wings held steeply tectiform in repose. This concavity serves for optimal position of the hind legs before jumping. Issidae have no such concavity, as their legs are covered by forewings before jumping and are angled differently at
jumping ( Anufriev & Emeljanov, 1988, fig. 11: 7). The forewing in Issidae has the bulla in basal half under R 1 close to first furcation of the radius. Most issid species have convex forewings except, for example, Oronoqua Fennah, 1947 and Eupilis Walker, 1857 with flat wings; however, these genera have forewings with convex costal margins, without a concavity. The forewing imprint of Uphodato garwoterus Szwedo, 2019 exhibits a distinct “cixiid clavus”, with Pcu + A 1 joining the wing margin before the claval apex, and with tubercles on setal bases on the veins ( Fig. 1 View FIGURES 1–4 ) which are characteristic features of Cixiidae , but not known for Achilidae .
Uphodato Szwedo, 2019 and Breukoscelis Szwedo, 2019 are distinguished by the stem R + M on the forewing ( Figs 1, 2 View FIGURES 1–4 ). Such a condition is known for extant Dilacreon orpheus ( Fennah, 1956) ( Fig. 4 View FIGURES 1–4 ), described from Caroline Islands ( Fennah, 1956), within the Eucarpiini. Uphodato and Breukoscelis are also characterized by a unique vein which may be treated as a transverse m-cua crossvein based on its position on the wing ( Figs 1–3 View FIGURES 1–4 ), but very likely it is the “root” of CuA, becoming a medio-cubital vein as a result of “interception” of the CuA by the M. In favor of this interpretation is the wing venation in members of the genus Dilacreon Fennah, 1980 illustrated by Fennah (1956, 1980), in particular, D. orpheus , which has a short stem R + M. However, on the drawings for Uphodato and Breukoscelis ( Szwedo et al., 2019, figs 46–48) CuA is not furcating, which is not typical for Cixiidae . If our interpretation of vein “interception” is true, then CuA acquires the usual character of furcating. Thus, the species of the genus Dilacreon have the basal part of CuA 1 and prenodal vein m-cua situated symmetrically as though ready for “interception”. The extravenal pterostigma is not visible on the imprints of fossil taxa; however, within extant Eucarpiini it is small or completely obsolete.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Fulgoroidea |
|
Family |