
Microlepidogaster negomata Martins, Cherobim, Andrade & Langeani, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4276.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:FEA7B464-F489-4397-B3D3-7E92CA358529 |
|
DOI |
https://doi.org/10.5281/zenodo.6045740 |
|
persistent identifier |
https://treatment.plazi.org/id/7A38FB6A-D06B-F63D-FF30-B7B5FCEF9DF9 |
|
treatment provided by |
Plazi |
|
scientific name |
Microlepidogaster negomata Martins, Cherobim, Andrade & Langeani |
| status |
sp. nov. |
Microlepidogaster negomata Martins, Cherobim, Andrade & Langeani , sp. n.
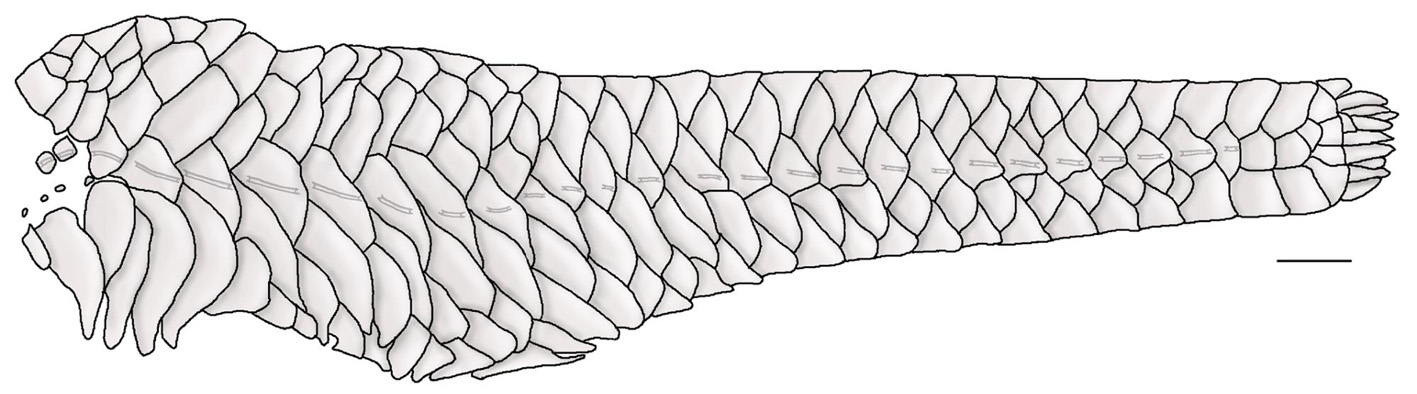
( Fig. 1 View FIGURE 1 ; Tab. 1)
Microlepidogaster sp. nov. —Martins et al. 2014: 827 (infraorbital series drawing), 847, 854, 856, 864, 865, 867, 868, 872 (phylogenetic relationships).
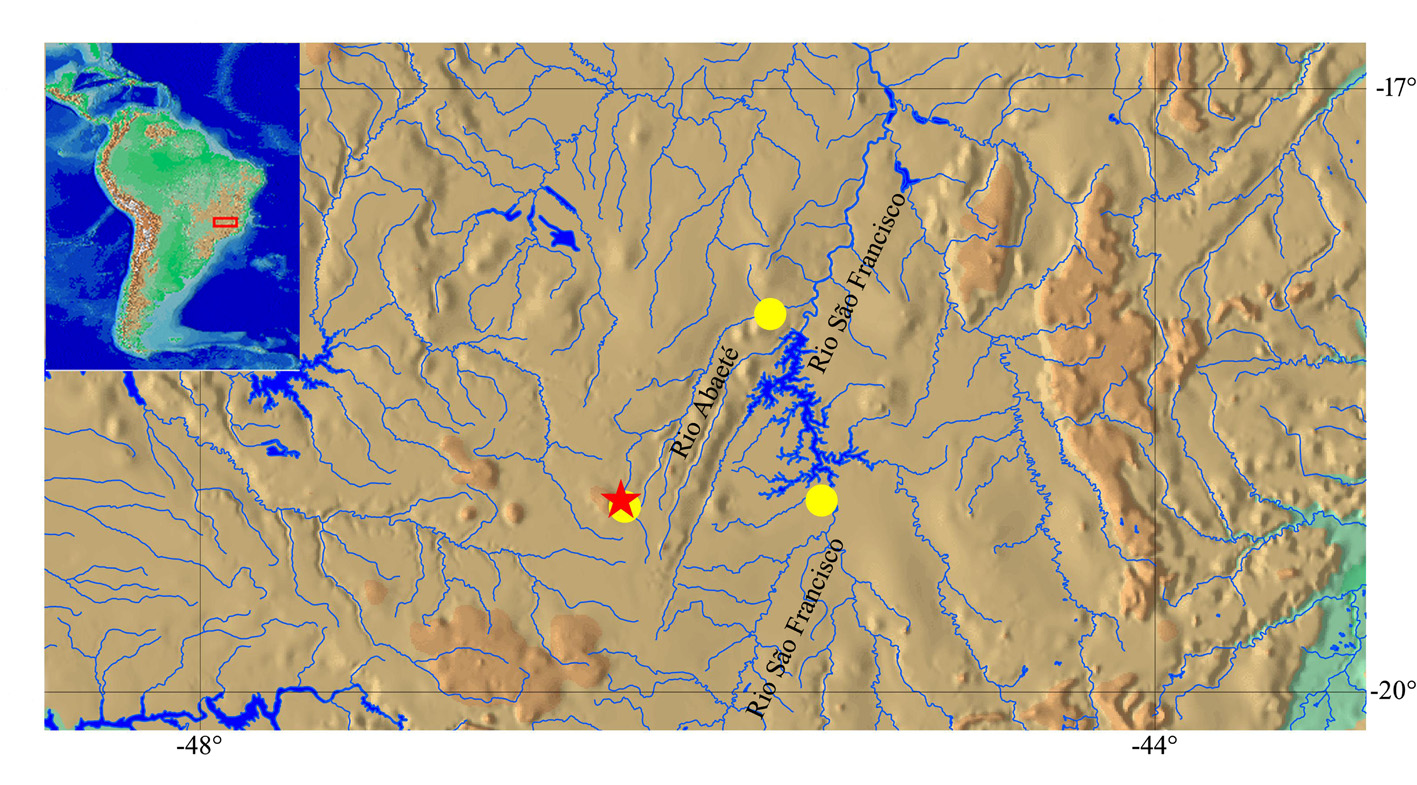
Type material. Holotype. DZSJRP 20135, male, 41.1 mm SL, Brazil, Minas Gerais State, Arapuá, Ribeirão Bebedouro on road to Chaves , upstream Arapuá , Rio Abaeté drainage, Rio São Francisco basin, 19°02’50”S 46°09’26”W, 920 m asl., 27 Aug 2014, F. Langeani, A.M. Cherobim, B.N. Andrade, A.C. Rosa. GoogleMaps
Paratypes. All from Brazil, Minas Gerais State, Rio São Francisco basin, Rio Abaeté drainage. DZSJRP 21003, 26 (1 c&s, and tissue of 3 specimens), 15.8–39.5 mm SL, collected with holotype. DZSJRP 20128, 18 (tissue of 2 specimens), 28.8–37.2 mm SL, Arapuá , Ribeirão Bebedouro , fazenda Nego Matá , 19°04’42”S 46°08’17”W, 1032 m asl., 27 Aug 2014, F. Langeani, A.M. Cherobim, B.N. Andrade, A.C. Rosa. LBP 11774, 2, 19.1–29.6 mm SL, Arapuá, 19°02’51”S 45°09’32”W, 19 May 2011, R. Devidé, G.J.C. Silva, R. Britzke. MZUSP 95291, 11 (1 c&s), 26.9–38.1 mm SL, Três Marias, Rio Curral das Éguas, on the border of the municipalities of Três Marias and São Gonçalo do Abaeté, 18°07’13”S 45°24’52”W, 3 Oct 2007, O. Oyakawa, M. Toledo-Piza, I. Fichberg, C. Chamon. GoogleMaps
Diagnosis. Microlepidogaster negomata differs from all congeners by having two to five unperforated platelets on skin covering swimbladder opening, anterior to the median lateral series of plates ( Fig. 2 View FIGURE 2 ) (vs. absence of these platelets in M. discontenta , M. discus , M. longicolla , and M. perforata ; and none or one platelet in M. arachas and M. dimorpha ); and ventral laminar expansion of coracoid enclosing totally or almost totally the arrector fossa of pectoral girdle ( Fig. 3 View FIGURE 3 E) (vs. ventral laminar expansion of coracoid enclosing partially the arrector fossa, Fig. 3 View FIGURE 3 A–D). In addition, M. negomata can be distinguished from all congeners, except for M. discus , by having the ventral laminar expansions of cleithrum and coracoid bearing 20–35 odontodes restricted to the lateral portion of pectoral girdle (vs. more than 35 odontodes, generally around 100, reaching the middle portion of pectoral girdle).
The following features are also useful to diagnose M. negomata within the genus: snout with many irregularshaped platelets, eventually with a naked area at the most anterior portion (vs. snout broadly naked anteriorly in M. discontenta and M. longicolla ; snout with paired rostral plate in M. dimorpha and M. perforata ; and snout with variable coverage of plates, from many platelets to a pair of rostral plates in M. arachas ); iris operculum present (vs. absent in M. perforata ); pectoral axillary slit present only in juveniles (vs. pectoral axillary slit persistent, in both juveniles and adults in M. dimorpha and M. perforata ); odontodes at anterior plates of snout small and slightly pointed (vs. leaf-shaped odontodes in M. dimorpha and M. perforata ); anterior portion of compound supraneural plus first dorsal-fin proximal radial contacting the neural spine of the eighth or ninth vertebra (vs. seventh vertebra in M. discus and M. dimorpha , tenth vertebra in M. discontenta , and tenth or eleventh vertebra in M. longicolla ); mid-dorsal lateral series of plates surpassing the vertical through the end of dorsal fin (vs. mid-dorsal lateral series reduced, never surpassing that point in M. longicolla and M. perforata ); median series of lateral plates complete, reaching the end of the caudal peduncle (vs. median series of lateral plates terminating two plates before the end of the caudal peduncle in M. perforata ); and first anal-fin pterygiophore covered by skin only, not bearing odontodes (vs. bearing odontodes in M. dimorpha and M. perforata ).
Description. Morphometric and meristic data given in Tables 1 and 2. Dorsal body profile slightly convex from tip of snout to tip of parieto-supraoccipital; almost straight to caudal-fin origin. Ventral body profile almost straight from tip of snout to pelvic-fin origin; ascending from pelvic-fin origin to end of anal-fin base; straight to caudal-fin origin. Greatest body depth variable, at parieto-supraoccipital tip or at dorsal-fin origin. Greatest body width at posterior margin of opercle, gradually tapering towards to caudal fin. Caudal peduncle ellipsoid in transverse section, slightly flattened dorsally and ventrally.
Anterior margin of snout rounded in dorsal view; tip of snout bearing many small irregular-shaped plates, or with naked areas at most anterior portion (more evident in juveniles). Eye small, dorsolaterally placed. Iris operculum present. Compound pterotic quadrangular in shape, its posterior extension poorly-developed, not reaching to rib of sixth vertebra; compound pterotic fenestrae irregular in shape and variable in size, smaller in dorsal and larger in ventral portion of bone. Infraorbital canal entering infraorbital series via sphenotic. Parietosupraoccipital not forming dorsal wall of swimbladder capsule.
Odontodes on head and body small and slightly pointed, not forming conspicuous rows. Bony crests and hypertrophied odontodes on parieto-supraoccipital and compound pterotic absent.
Lips oval, papillose; lower lip not reaching pectoral girdle; papillae gradually smaller toward lip edges. Maxillary barbel reduced, free from oral disk. Teeth slender and bifid; median cusp larger and rounded, lateral cusp smaller and pointed. Premaxillary teeth 16–26 (19). Dentary teeth 14–25 (22). Accessory teeth present only in juveniles.
Dorsal-fin rays II,6–7; originating approximately at vertical through end of pelvic-fin base; tip of adpressed rays almost reaching vertical through end of anal-fin base; spinelet small, somewhat elliptical in shape, locking mechanism non-functional. Anterior portion of compound supraneural plus first dorsal-fin proximal radial contacting neural spine of eighth or ninth vertebra. Supraneural anterior process absent. Pectoral-fin rays I,6; originating immediately behind opercular opening; tip of adpressed rays reaching to vertical through end of pelvicfin base. Pectoral-fin unbranched ray without ossified segments distally. Cleithrum and coracoid only exposed laterally, restricted to area near to pectoral-fin insertion, with lower number of odontodes (20–35) compared to congeners; remaining portions of pectoral girdle covered by skin and abdominal plates. Arrector fossa totally or almost totally enclosed. Pectoral axillary slit present only in juveniles. Pelvic-fin rays i,5; unbranched ray shorter than branched rays. Anal-fin rays i,5. Caudal-fin rays i, 13-15(14), 1; concave; lobes equal in size; 3–5 (4) dorsal and 2–5 (3) ventral procurrent rays. Adipose fin absent; some specimens with one or two rounded or elongate azygous plates in place of adipose fin.
Body entirely covered by dermal plates, except for ventral part of head, region overlying opening of swimbladder capsule, around pelvic-fin origin, and region anterior urogenital opening. Abdomen entirely covered with small-sized plates irregularly distributed; lateral abdominal plate series absent. Mid-dorsal series of lateral plates surpassing vertical through end of dorsal fin. Median lateral plate series complete and uninterrupted with 27– 31 (29) plates. Two to five reduced unperforated platelets on skin covering swimbladder opening, anterior to median series of plates. Vertebrae 32 (32).
Ventral lateral plates 22(3), 23(5), 24(3), 25(12), 26(11)*, 27(3), 28(2), 29(6), 30(1) 22–30 25
Premaxillary teeth 16(1), 17(0), 18(4), 19(10), 20(6), 21(1), 22(5), 23(6)*, 24(4), 16–26 19
25(1), 26(1)
Color in alcohol. Ground color of head and body light-brown on dorsal, lateral and ventral portions. Head dark brown along parieto-supraoccipital and area from anterior border of snout to eye. Rectangular, longitudinally elongate, dark-brown blotch between compound pterotic and posterior portion of cleithrum process and plates at mid-ventral series. Dark brown lateral stripe inconspicuous in most specimens, approximately two plates wide, extending from anterior portion of snout and fading on caudal peduncle. Four dorsal dark brown blotches, conventionally arranged as other congeners; more conspicuous in melanic specimens. Pectoral, dorsal, pelvic, and anal fins mostly hyaline, with tiny light-brown chromatophores along rays. Caudal fin almost entirely dark brown, except for distal tip of rays and a hyaline blotch in each lobe; greater specimens with blotches inconspicuous, darkened.
Sexual dimorphism. Males with conspicuous urogenital papilla immediately posterior to anus (vs. absent in females) and also with a dermal flap on dorsal surface of unbranched pelvic ray and first branched pelvic-fin rays (vs. absent in females). Males with longer pelvic-fin extending to or surpassing anal-fin origin (vs. pelvic-fin never reaching anal-fin origin in females).
Distribution. Microlepidogaster negomata is known from the Ribeirão Bebedouro and Rio Curral das Éguas, Rio Abaeté drainage, tributary to left side of the Rio São Francisco, Minas Gerais state, southeastern Brazil ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ).
Ecological notes. The type locality of Microlepidogaster negomata ( Fig. 5 View FIGURE 5 ) is a very small stream near the Arapuá municipality, in a stretch between 912 to 1032 meters above sea level. The stream is about 0.8 m wide, up to 0.4 m deep, with bottom composed of gravel, stones, and sand. The riparian vegetation is mainly composed of grasses and herbs and surrounded by a grazing area with a few palms and shrubs, which favors bank erosion and great penetration of light. Microlepidogaster negomata was collected mainly associated with marginal vegetation and other species collected in the same stream include Astyanax bimaculatus ( Linnaeus, 1758) (DZSJRP 20137), A. rivularis ( Lütken, 1875) (DZSJRP 20126), Cetopsorhamdia iheringi Schubart & Gomes, 1959 (DZSJRP 20131), Harttia sp. (DZSJRP 20129), Imparfinis sp. (DZSJRP 20133), Parotocinclus prata (DZSJRP 20130), Trichomycterus sp. 1 (DZSJRP 20136), and Trichomycterus sp. 2 (DZSJRP 20132).
Etymology. The specific epithet, negomata , is in reference of a locality in Ribeirão Bebedouro where the species was captured, the Fazenda Nego Matá. The denomination, Nego Matá, was the nickname of the former owner of the farm, Mr. Esli Domingues. A noun in apposition.



| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |