
Paranaitis
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00069.x |
|
persistent identifier |
https://treatment.plazi.org/id/793A8787-5C0A-601B-FC68-FBB0FBB7CC03 |
|
treatment provided by |
Carolina |
|
scientific name |
Paranaitis |
| status |
|
MORPHOLOGY OF PARANAITIS View in CoL
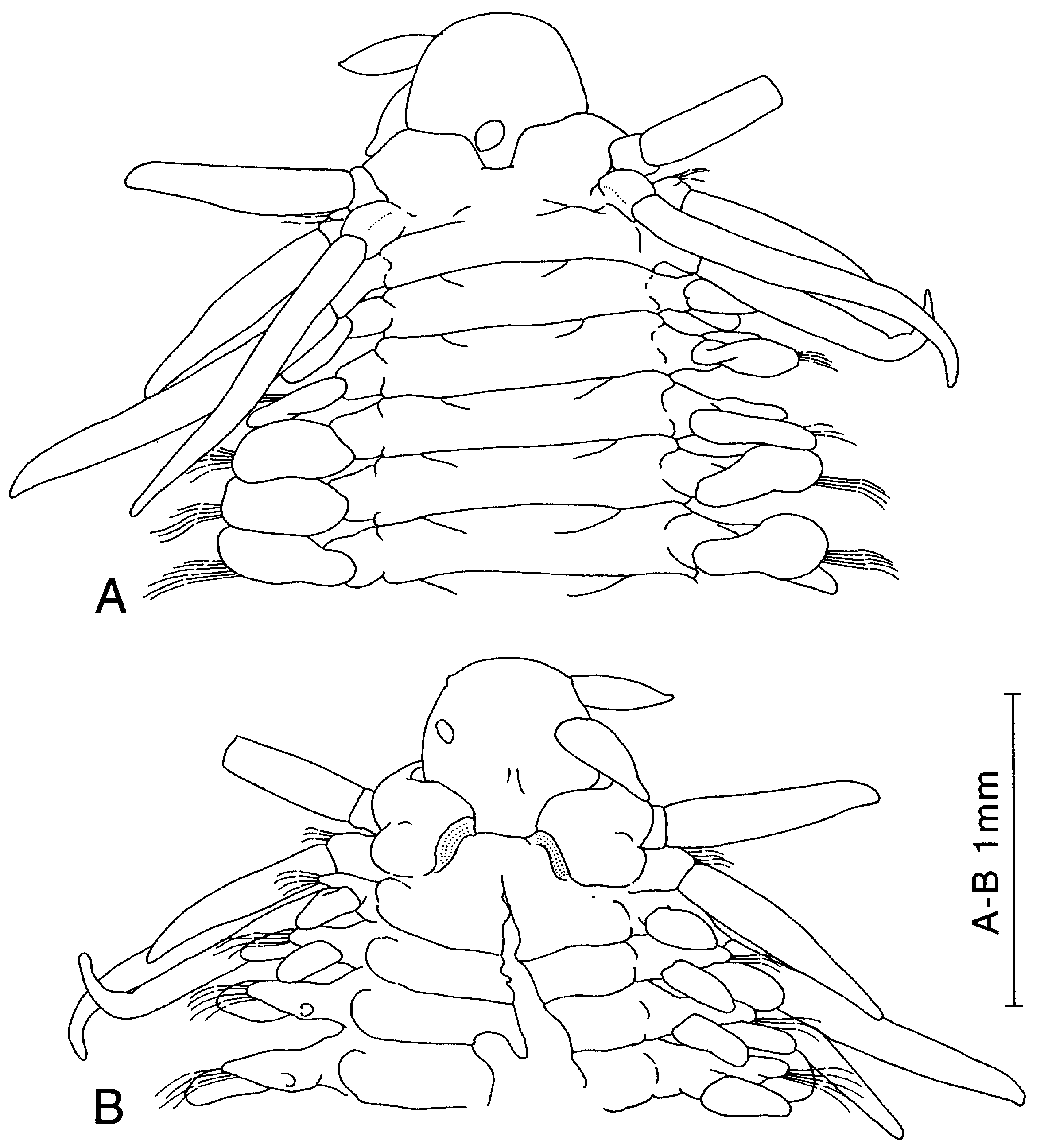
The shape of the prostomium may be rounded to triangular, although the differences between taxa are gradual and poorly defined. The posterior part of the prostomium is always covered by segment 1. There are two pairs of front appendages of which the dorsal pair is labelled ‘paired antennae’ and the ventral pair ‘palps’ (homologous to palps in other Phyllodocida , e.g. Orrhage & Eibye-Jacobsen, 1998). The paired antennae and the palps may be antero-laterally orientated and inserted on a rounded to flat prostomial surface ( Fig. 5A, B View Figure 5 ), or they may be postero-laterally orientated and inserted in lateral prostomial depressions ( Fig. 13A View Figure 13 ). This character is potentially informative, although there are difficulties in discriminating states in some specimens, especially when they are in a poor condition. A single pair of eyes with lenses may be present, or eyes are entirely absent. A single nuchal papilla is situated near the posterior margin of the prostomium in an incision (ligula) surrounded by segment 1, and can be indistinct (e.g. P. wahlbergi ), short and rounded (e.g. P. kosteriensis ), or long and pointed and similar to the paired antennae and the palps (e.g. P. caeca ). The ligula may be deep or shallow.
The proboscis morphology is highly variable in Paranaitis and provides a series of characters. It can be subdivided into distal and proximal parts (referring to the proboscis in everted state) or not subdivided; in the subdivided proboscis the parts can be sharply delineated or have a gradual transition. In taxa with a subdivided proboscis the distal part is usually covered by six rows of large rounded tubercles, of which the two dorsal rows can merge into a single one. The proximal part lacks tubercles, but is covered by minute chitinous papillae which may also be present on the distal part. These papillae are very small and transparent, and their detection requires examination with a compound microscope. Furthermore, under the epithelium of the proboscis, there are one or a few kinds of rounded structures ( Fig. 17B–E View Figure 17 ) which are possibly secretory glands, and which have frequently been misinterpreted as papillae (e.g. Pleijel, 1993a for Paranaitis near polynoides ). The size and shape of the chitinous papillae are quite variable and related to their position on the proboscis. A group of large papillae may be present in transverse rows on each side of the proximal-most part. In taxa with a nonsubdivided proboscis, the surface may be smooth or covered by tubercles and large or small papillae. A longitudinal lateral row of large papillae may be present on each side.
The nuchal organs in Paranaitis have not been detailed in the literature. They are retractile, with horseshoe-shaped bands of cilia, but vary in size and position between species. The nuchal organs can be completely retracted and invisible, and can also be difficult to examine in specimens in poor condition.
Segments 1 and 2 are large, more or less fused to each other, and form a collar around the prostomium. The degree of fusion of these two segments has been thought to be diagnostic for certain species (e.g. P. polynoides ). Nevertheless, the interpretation of this character is problematic due to the presence of fixation and preservation artefacts.
The cirri of segment 1, the dorsal and ventral cirri of segment 2 and the dorsal cirri of segment 3 differ in shape from the following dorsal and ventral cirri in being long and digitiform rather than rounded and flattened, and are traditionally labelled ‘tentacular cirri’. Although the term is unproblematic within the Phyllodocidae , it is equivocal for a number of other polychaete groups within Phyllodocida (see e.g. Pleijel, 1998), and conflates the actual shape of cirri with other characters, such as a reduction of chaetae and chaetigerous lobes on the anterior segments. Referring to these cirri with a specification of their position (e.g. dorsal cirri of segment 2) and shape (e.g. digitate) increases the precision of the descriptions and eases comparisons between more distant polychaete taxa. For this reason, we avoid here the use of the term ‘tentacular cirri’. The length of the anterior cirri exhibits interspecific differences, although considerable intraspecific variation is present in P. wahlbergi . The presence or absence of dorsal aciculae in segments 2 and 3 is potentially informative among Paranaitis , although unknown for some rare species (e.g. P. abyssalis ). The chaetae may be present on segment 2, arising from neuropodial lobes or from the cirrophores of the ventral cirri. From segment 3 and onwards they always emerge from the neuropodial lobes. The number of chaetae varies between species (although not in a very fixed pattern) and is also related to body size. Segment 4 and the following segments are provided with uniramous parapodia with reniform flattened dorsal and ventral cirri. The outline of the dorsal cirri is oval, circular or broadly reniform. The dorsal cirrophores show little intraspecific variation, and are large and distinct in some species and short and indistinct in others. In P. uschakovi they carry a ciliary band. The dorsal cirrophores may be well developed or indistinct, and when well developed they can be symmetrical around the longitudinal axis ( Fig. 13C–F View Figure 13 ), they can be bilobed with enlarged dorsal lobes ( Fig. 8F–I View Figure 8 ), or they can be thin with dorsal extensions ( Fig. 22H–I View Figure 22 ). The supra-acicular lobes of the neuropodium are longer than the subacicular lobes. The ventral cirri have rounded-to-pointed ends, with the longitudinal axis orientated horizontally. All chaetae are compound in Paranaitis . The rostrum of the chaetal shaft is covered by a large number of small teeth, and may have a single to a few additional main teeth on the anterior side, or both on the anterior and posterior sides. When the anterior and posterior sides are similar to each other, the rostrum is labelled as symmetrical; when different it is labelled as asymmetrical. The shape of the chaetae is similar from the dorsal to the ventral part of the fascicles in most species, although gradual changes from asymmetrical to symmetrical are found in P. inflata and P. kosteriensis . The chaetal blades are always long and slender, and we could not detect any informative variation in them (see also remarks for P. inflata ).
A pair of pygidial cirri and a single pygidial papilla are present, although unknown for some species. The pygidial cirri may be rounded or cylindrical, and have rounded-to-pointed ends.
The colour and pigmentation pattern of live Paranaitis , as in many other phyllodocids, exhibits important interspecific differences. Many species have whitish bodies with a red pigmentation that is unusual (possibly unique except some species of Mysta ) in phyllodocids, although other pigmentation patterns are also present. In some taxa the pattern may be retained in well-preserved specimens and provide additional information for identification; in others it disappears completely.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.