
Singhiella melanolepis Chen & Ko
|
publication ID |
https://doi.org/ 10.5281/zenodo.175204 |
|
DOI |
https://doi.org/10.5281/zenodo.5667907 |
|
persistent identifier |
https://treatment.plazi.org/id/790A87D4-FF82-2830-FF36-FC49591E2D8D |
|
treatment provided by |
Plazi |
|
scientific name |
Singhiella melanolepis Chen & Ko |
| status |
sp. nov. |
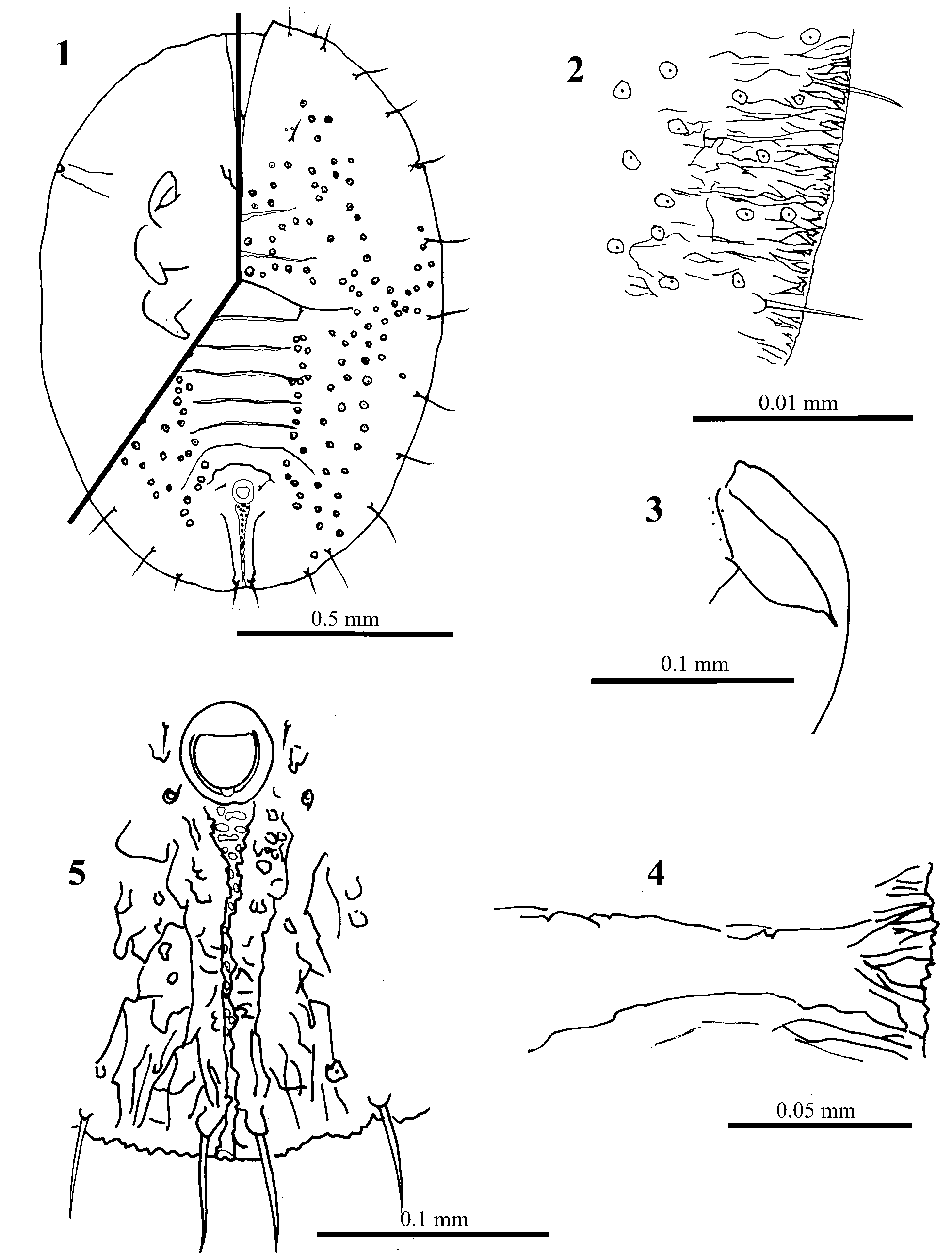
Singhiella melanolepis Chen & Ko View in CoL sp. nov. ( Figs 1–36 View FIGURES 1 – 5 View FIGURES 6 – 9 View FIGURES 10 – 11 View FIGURES 12 – 18 View FIGURES 19 – 27 View FIGURES 28 – 34 View FIGURES 35 – 36 )
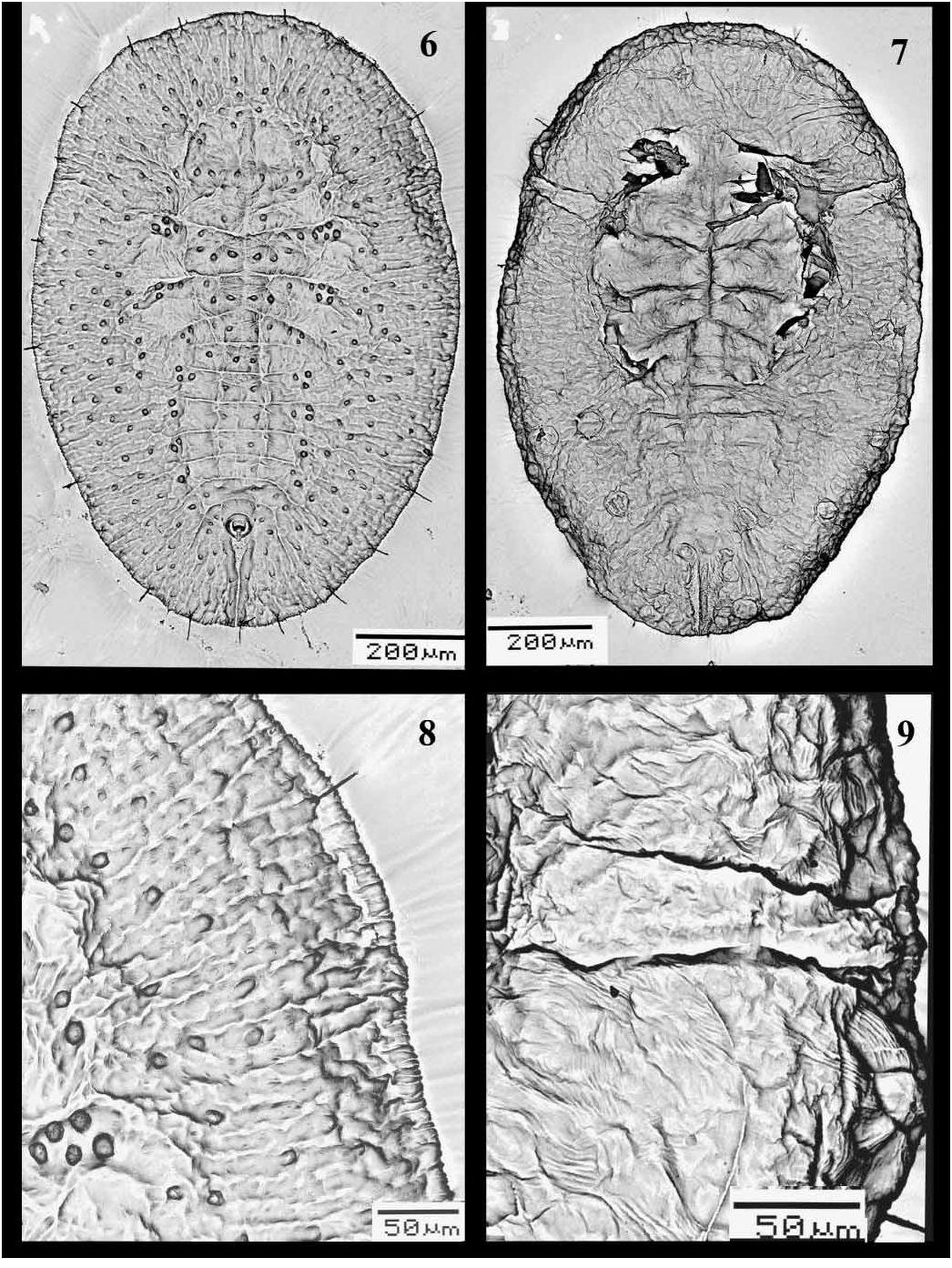
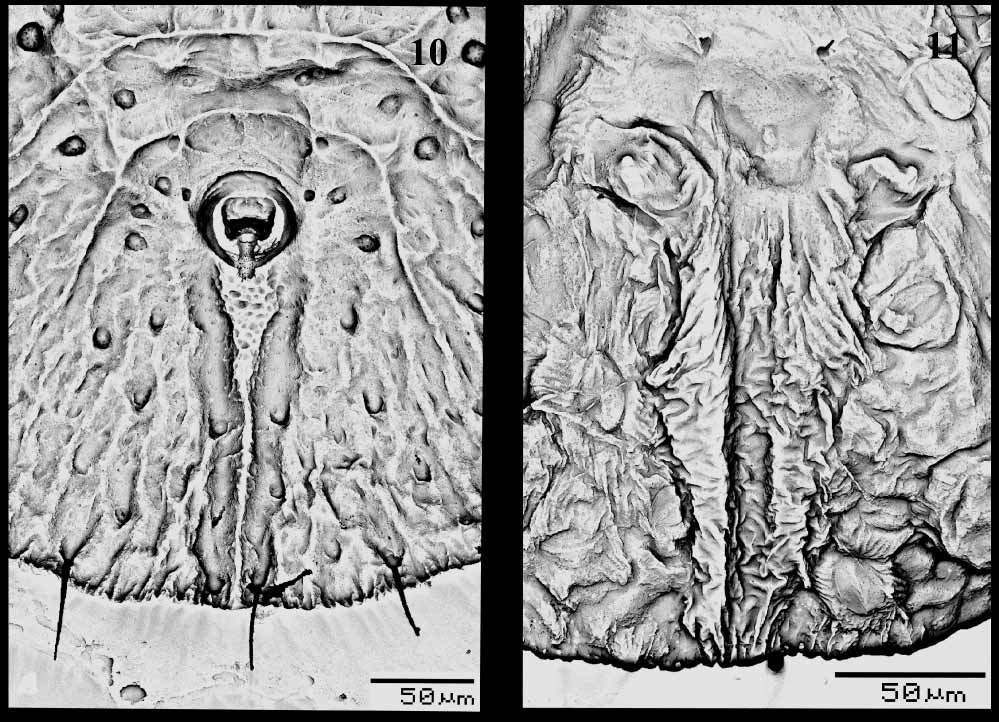
PUPARIUM ( Figs 1–11 View FIGURES 1 – 5 View FIGURES 6 – 9 View FIGURES 10 – 11 ). Found in groups on the undersurface of leaves. Pale to yellowish, but generally colourless when slidemounted. Most pupae without, or with only a little, waxy secretion. Pupal case average 1.35 mm long, 1.05 mm wide, elongate to ovoid, broadest at first abdominal segment. Margin more or less smooth, or with faint crenulations marked by short radiating ridges, about 24 ridges per 100 μm. Ends of thoracic and caudal tracheal folds not evident and marked by short smooth areas in margin ( Fig. 2 View FIGURES 1 – 5 ). Dorsum. Moreorless flat, with raised rachis or other ridges, usually on cephalothorax and dorsal disc, abdominal tergites I–VII without median tubercle but subdorsal tubercles on cephalothorax and on each side of tergites III–VI. Sculpturing of fine papillae and inscriptions, in subdorsal areas arranged in less radiating rows of papillae; sculpturing sometimes highly reduced. Longitudinal moulting suture more or less straight, reaching margin. Transverse moulting suture reaching a point just above or slightly beyond lateral margins of hind legs; ends of suture with distinct raised areas or clusters of papillae, slightly posterior relative to middle of suture. Eighth abdominal segment subequal to seventh in length. Disc pores and associated porettes scattered over dorsum; each pore/porette pair not close together, pores uniformly mounted on tubercles or papillae but porette not. Setae. Marginal setae pointed, on anterior and posterior margins, anterior pair 0.02 mm long, posterior pair 0.045 mm long. Twelve pairs of submarginal setae ( Figs 2 View FIGURES 1 – 5 , 8 View FIGURES 6 – 9 ) of nearly uniform length, 0.05 mm; sixth submarginal setae usually in line with other submarginal setae and not inset medially to nearly above middle leg; twelfth pair of submarginal setae not inset on caudal ridges. Cephalic setae 0.025 mm long, distinctly forward of mouthparts. First abdominal setae present. Eighth abdominal setae laterad of vasiform orifice, more or less in line with anterior margin. Caudal setae slightly anterior to margin, 0.5 mm long. Vasiform orifice ( Figs 5 View FIGURES 1 – 5 , 10 View FIGURES 10 – 11 ). More or less trapezoidal to almost circular, 0.06 mm long and 0.055 mm wide, middle of posterior margin not broken, often with loose tilelike structures in notched area; inner margin often with faint ridges radiating into orifice; operculum equal in size to vasiform orifice and similar in shape, nearly filling orifice, usually without posterior part distinctly narrowed; lingula exposed, without setae, almost included, except a small part of posterior end exposed outside. Caudal furrow ( Figs 5 View FIGURES 1 – 5 , 10 View FIGURES 10 – 11 ). With a larger tilelike structure, 0.225 mm long, less wide than vasiform orifice anteriorly, abruptly narrowed posteriorly, reaching margin; furrow lined with large granules in more or less transverse rows on wide part, more scattered on narrow part. Caudal ridges apparently slightly raised above adjacent derm, especially posteriorly. Area lateral to vasiform orifice and caudal furrow without spinules.
Ventral surface ( Figs 4 View FIGURES 1 – 5 , 7, 9 View FIGURES 6 – 9 , 11 View FIGURES 10 – 11 ). Thoracic and caudal tracheal folds usually distinguishable but entirely smooth; caudal fold with a few short rows of minute spinules extending about half way to margin from vasiform orifice. Ventral setae finely pointed, posterior pair just anterior to vasiform orifice. One small, fine seta mesad of each leg, normally 2 minute setae on or near basal curve of each leg, often difficult to see. Legs curved laterally, front and middle legs on each side closely apprised; posterolateral margin of front legs and lateral margins of middle and hind legs without spinules. Antenna located just mesad of front leg, 0.07 mm, terminal process 7.5 μm, lacking spinules at base, with a minute seta basally. Rostrum short, conical. Four pairs of spiracles present, 2 near posterior part of front and middle legs, one just beyond hind legs on anterior abdomen and one just laterad of vasiform orifice.
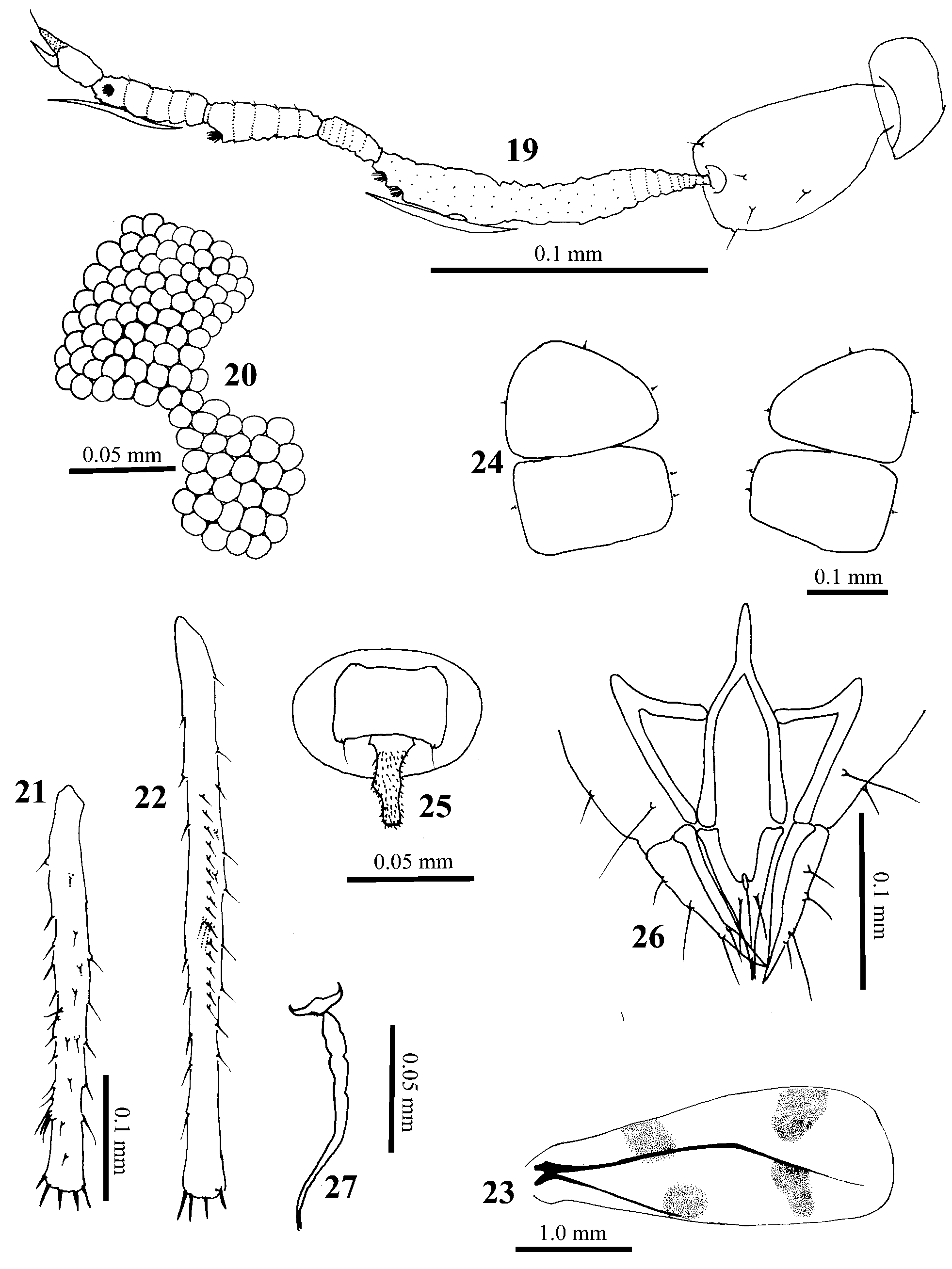
ADULT MALE ( Figs 12–18 View FIGURES 12 – 18 , 28, 32–34 View FIGURES 28 – 34 ). Yellowish to brown when alive, with a light dusting of wax. Middle and distal end of forewing with four dusky spots. Yellowish when mounted, except extreme tip of rostrum and thorax brown. Average body length 1.5 mm (included claspers). Essentially entire body covered with minute setae. Head. With scattered minute setae with short cylindrical bases. Width across eyes 0.3 mm. Rostral IV 0.125 mm measured along longest edge, with 15–20 setae. Antennae ( Figs 12 View FIGURES 12 – 18 , 28 View FIGURES 28 – 34 ). Inserted in median indentation of compound eyes; segment I less than half as long as II; segment II with several scattered setae of various sizes; segment III 0.125 mm, with 2 primary sensoria apically, one about its width basad of other, one bidirectional sensorial cone more basad 0.045 mm; segment IV 0.02 mm; V 0.045 mm, with one primary sensorium apically; VI 0.03 mm, with one bidirectional sensorial cone near middle, about as long as sensorial cone on segment III; VII 0.0325 mm, with one primary sensorium a little beyond middle of segment and one sensorial cone 0.015 mm in basal third of segment, apical spine 0.02 mm; apparent annulations of antenna composed of very closeset setulae. Compound eyes ( Figs 13 View FIGURES 12 – 18 , 32 View FIGURES 28 – 34 ). Constricted laterally, dorsal and ventral parts connected by 2 facets; ventral facets about 50% larger than dorsal facets. Lateral ocelli abut dorsal margin of compound eyes, about twice as wide as dorsal eye facets. Thorax. Sclerotized plates typical for aleyrodine adults. Wings with a patch of maculation in middle and near distal end. Legs. Coxa 1, 0.145 mm; femur 1, 0.2 mm, femur 2, 0.2 mm, femur 3, 0.263 mm, including dorsal toothlike projection at extreme apices, femur 3 with 3 strong subapical spines dorsally; tibia 1, 0.263 mm, with about 45 setae and no tibial brushes, tibia 2 ( Fig. 14 View FIGURES 12 – 18 ), 0.3 mm with 1 tibial brush with 2 setae and sometimes a third seta close but slightly basad, tibia 3 ( Fig. 15 View FIGURES 12 – 18 ), 0.385 mm with about 12 setae in tibial comb, without tibial brush, and with a distinct nonsetose area basad of apical spines; tarsal segment I with following lengths and setae (tarsi 1, 2, 3): 0.105 mm with 6–8 setae, 0.105 mm with 8–10 setae, 0.12 mm with 7–9 setae; tarsal segments II with following lengths and setae (tarsi 1, 2, 3): 0.0875 mm with 7–8 setae, 0.08 mm with 4 setae, 0.0875 mm with 4 or 5 setae; hind tarsal claw 0.0375 mm. Abdomen. Ventral surface with 4 pairs of wax plates on segments II–V ( Fig. 17 View FIGURES 12 – 18 ), length/width of plate on segment II = 0.08 mm / 0.12 mm, III = 0.0575 mm / 0.13 mm, IV = 0.0575 mm / 0.125 mm, V = 0.053 mm / 0.10 mm. Each wax plate with 2 pairs of setae inward but 1 pair outward. Genital capsule ( Fig. 18 View FIGURES 12 – 18 ). 0.113 mm long and 0.165 mm wide. Vasiform orifice ( Fig. 18 View FIGURES 12 – 18 ). Oval, wider than long, lingula 0.002 mm; capsule with 5–6 pairs of dorsal setae, lateralmost setae distinctly thinner and shorter than more medial pairs, most medial pair thinner than thick middle pair. Clasper ( Figs 18 View FIGURES 12 – 18 , 33–34 View FIGURES 28 – 34 ). 0.1 mm, with five pairs of dorsal setae, and four and three pairs of setae present on inner and outer surfaces respectively, small rounded tooth on medial surface subapically. Aedeagus 0.1 mm, curved up apically and forked at end.
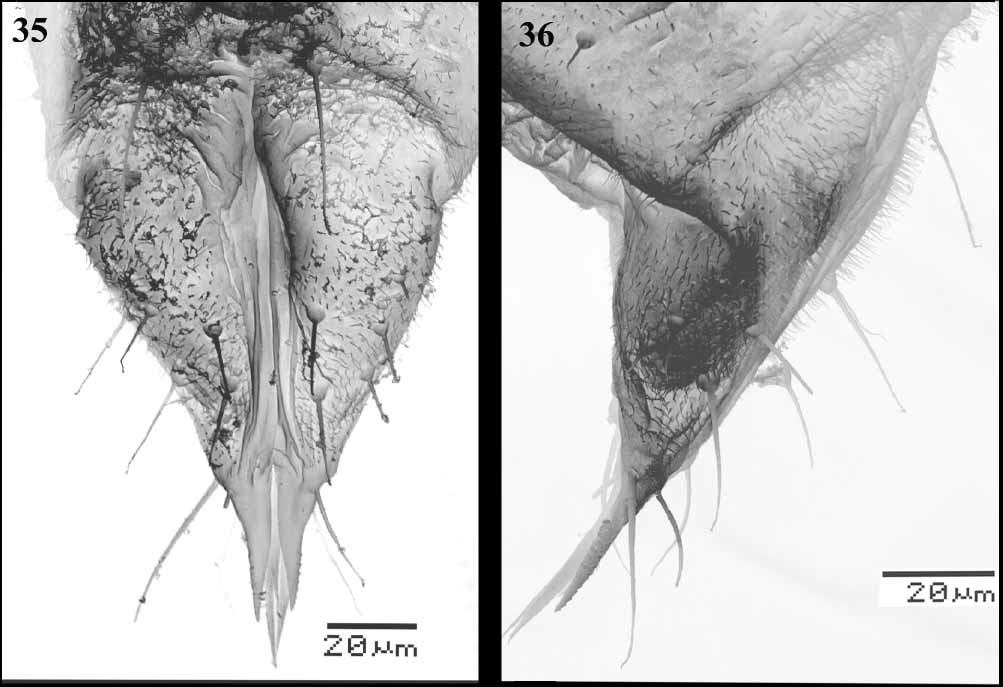
ADULT FEMALE ( Figs 19–27 View FIGURES 19 – 27 , 29 View FIGURES 28 – 34 , 35–36 View FIGURES 35 – 36 ). Colour as in male. Body length 2 mm. Head. Width across eyes 0.4 mm. Antennae. ( Figs 19 View FIGURES 19 – 27 , 29 View FIGURES 28 – 34 ). Inserted in median indentation of compound eyes; segment I less than half as long as II, segment II with several scattered setae of various sizes; III 0.138 mm, with 2 primary sensoria apically, one just distad of other and one bidirectional sensorial cone more basad 0.06 mm; IV 0.025 mm; V 0.043 mm, with one primary sensorium apically; VI 0.045 mm, with one bidirectional sensorial cone near middle, about as long as sensorial cone on segment III; VII 0.04 mm, with one primary sensorium a little beyond middle of segment and one sensorial cone 0.05 mm on basal third of segment, apical spine 0.015 mm. Rostral IV 0.135 mm measured along longest edge, with 15–20 setae. Compound eyes ( Fig. 20 View FIGURES 19 – 27 ). Constricted laterally, dorsal and ventral parts connected by 2 facets. Legs. Coxa 1, 0.163 mm; femur 1, 0.25 mm, femur 2, 0.24 mm, femur 3, 0.30 mm with a strong subapical spine dorsally; tibia 1, 0.14 mm, tibia 2 ( Fig. 21 View FIGURES 19 – 27 ), 0.16 mm, tibia 3 ( Fig. 22 View FIGURES 19 – 27 ), 0.22 mm with about 16 setae in tibial comb and with a brush with 3 setae; tarsal segment I with following lengths and setae (tarsi 1, 2, 3): 0.11 mm with 10 setae and a strong subapical spur, 0.1 mm with 8–9 setae, 0.125 mm with 6 setae; tarsal segment II with following lengths and setae (tarsi 1, 2, 3): 0.10 mm with 7 or 8 setae, 0.09 mm with 7 setae, 0.10 mm with 4 setae; hind tarsal claws 0.025 mm, paronychium 0.04 mm. Abdomen. Ventral surface with 2 pairs of wax plates ( Fig. 24 View FIGURES 19 – 27 ) on segments II and III, length/width of plate on segment II = 10.118 mm / 0.175 mm, III = 0.113 mm / 0.180 mm. Each wax plate with 2 inner but 1 outer pairs of setae. Vasiform orifice ( Fig. 25 View FIGURES 19 – 27 ). With lingula 0.03 mm. Genitalia ( Figs 26–27 View FIGURES 19 – 27 , 35–36 View FIGURES 35 – 36 ). With lateral ovipositor 0.095 mm, measured as shown in Fig. 26 View FIGURES 19 – 27 , with 4 setae; middle ovipositor 74–84 mm, with 2 setae. The cement gland slightly segmented ( Fig. 27 View FIGURES 19 – 27 ).
Material examined. Holotype puparium, TAIWAN, Chiaochi, on Melanolepis multiglandulosa ( Euphorbiaceae ), 22III2006, C. H. Chen and Y. T. Shih (TW 2961) ( NTU). Paratypes: 26 pupal cases, 7 males, 8 females, same data as for holotype ( ANIC; BMNH; EMNAFU; IDAV; NTU; TARI; USNM; ZSI).
Etymology. The species name melanolepis is derived from the host plant genus.
Biology. This species has been collected only once, and is of no known economic importance. No ant attendance was observed, but mixed populations of Aleurodicus dispersus Russell were observed on the lower leaf surfaces. Based on the distribution of Melanolepis multiglandulosa ( Euphorbiaceae ), this whitefly species is likely to have a wider distribution. It is a perennial shrub widely distributed in the South Pacific Islands from Indonesia to eastern Polynesia, also in China, Taiwan, and Japan. It grows mainly in secondary places such as roadsides, regrowth thickets, depleted open secondary forest, forest edges in savannah, coconut plantations, old gardens, but also in primary forest, ( Barringtonia ) swamp forest, Eucalyptus deglupta dominated forest, monsoon (deciduous) forest, and among mangroves.
Comments and discussion. In Taiwan, nine species are currently included in Singhiella : S. chitinosa (Takahashi) , S. dioscoreae (Takahashi) , S. elaeagni (Takahashi) , S. kuraruensis (Takahashi) , S. melanolepis sp. nov., S. piperis (Takahashi) , S. subrotunda (Takahashi) , S. tetrastigmae (Takahashi) and S. vanieriae (Takahashi) . S. melanolepis is similar to S. elaeagni but differs as follows:
1. All setae on the dorsal disc and all submarginal setae are capitate in S. elaeagni but spiny in S. melanolepis ;
2. There are 14 pairs of submarginal setae (excluding caudal setae) in S. elaeagni but only 12 pairs in S. melanolepis ;
3. Thoracic tracheal pores are distinct in S. elaeagni , with 2 or 3 small teeth, but there are no distinct pores in S. melanolepis ;
4. The vasiform orifice of S. elaeagni is wider than long and the end of the caudal furrow is widely open, with 2 or 3 very short rounded teeth, but such characteristics are not present in S. melanolepis .
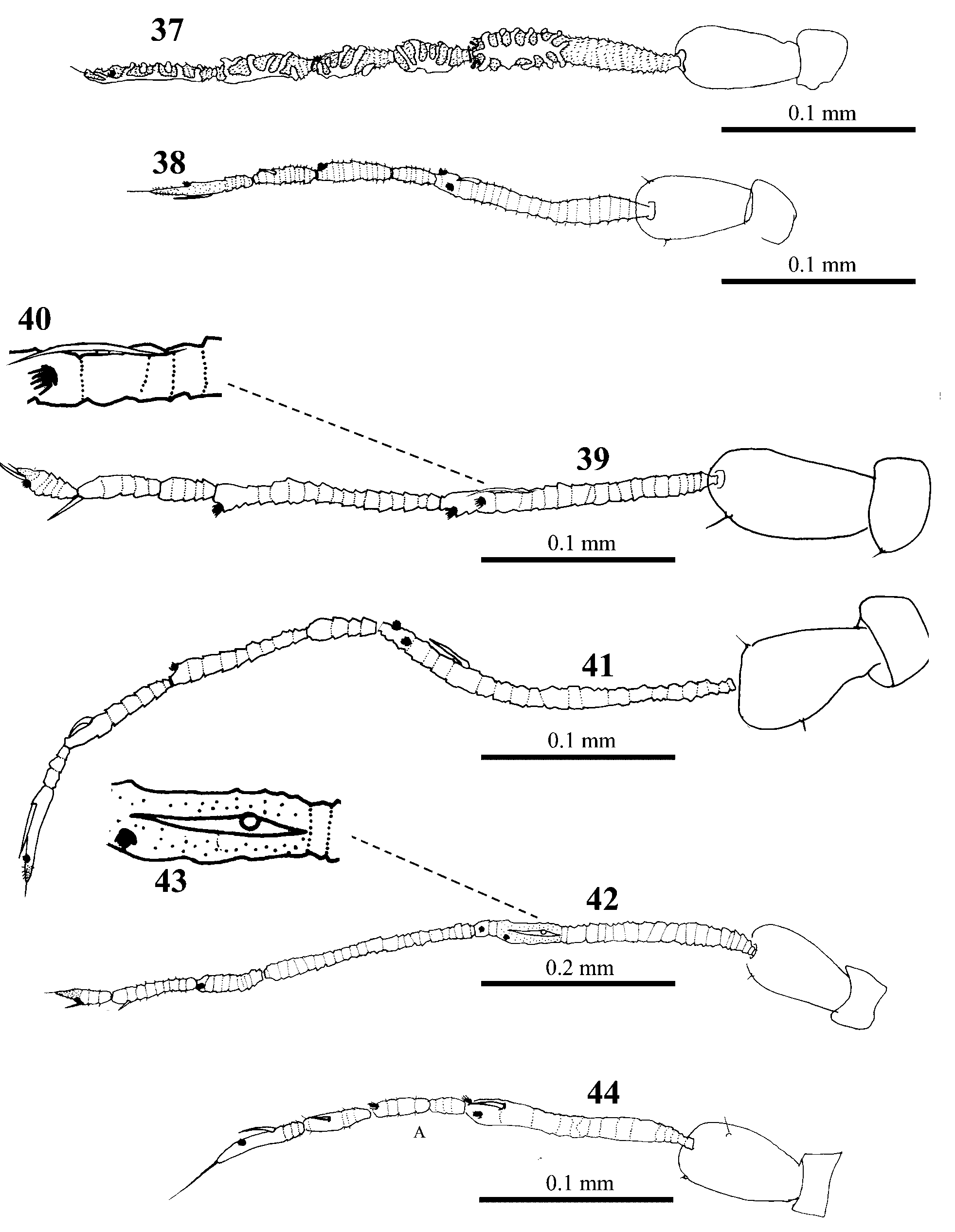
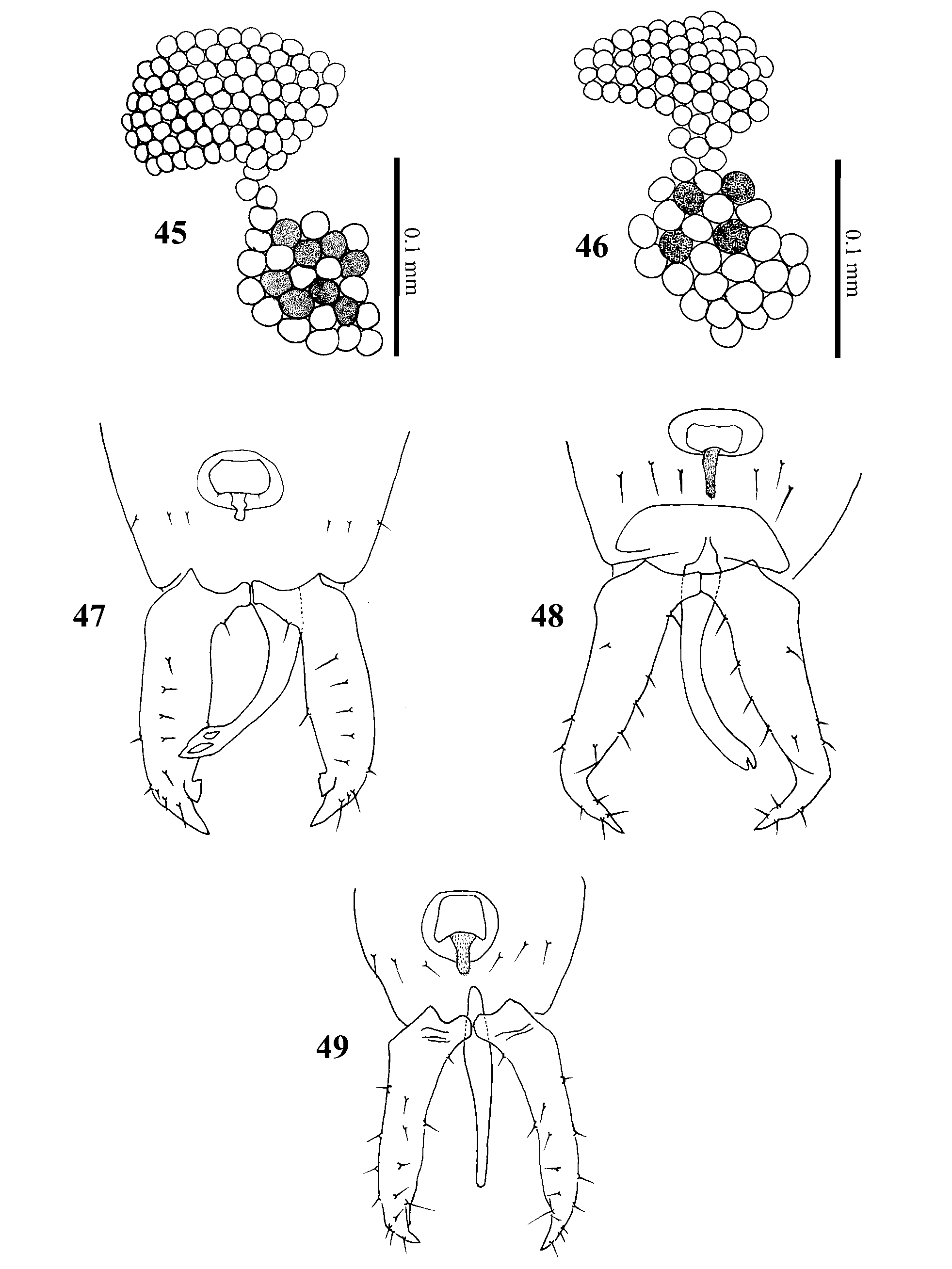
Following the redefinition of Singhiella by Jensen (2001), there are now 28 species in the genus. However, Singhiella is less clearly defined than Dialeurodes and Massilieurodes , with considerable variation in the puparia between species (see Table 2), including the opening of the thoracic tracheal pores, the number and shape of the submarginal setae, and the location of the most posterior pair of submarginal setae. Jensen (2001) described the adult characteristics of only one Singhiella species, S. citrifolli . In contrast, we have studied the adults of several species in this genus from Taiwan, and these show considerable variation in several characters. Antennal sensoria: We compared the adults of five species of Dialeurodes , five species of Massilieurodes and five species of Singhiella . The antennal segments of the Dialeurodes species bore complex sensorial plaques ( Fig. 37 View FIGURES 37 – 44 ), whereas those of the Massilieurode s species bore simple sensorial cones ( Fig. 38 View FIGURES 37 – 44 ). However, in different Singhiella species two types of sensorial cones (bidirectional and simple sensorial cones) were found on antennal segments III and VI ( Figs 39–44 View FIGURES 37 – 44 ). Compound eye pigmentation patterns: In species of Dialeurodes and Massilieurode s, we could not find pigmentation in the lower facets of the compound eyes, but in Singhiella we found pigmentation in two species, S. kuraruensis ( Fig. 45 View FIGURES 45 – 49 ) and S. piperis ( Fig. 46 View FIGURES 45 – 49 ). Male genitalia: In species of Dialeurodes and Massilieurode s, we did not observe any bifurcation in the aedeagus apex, but in Singhiella we found four types of aedeagus apex: S. kuraruensis ( Fig. 47 View FIGURES 45 – 49 ), S. melanolepis ( Figs 33–34 View FIGURES 28 – 34 ), S. piperis ( Fig. 48 View FIGURES 45 – 49 ), and S. tetrastigmae ( Fig. 49 View FIGURES 45 – 49 ). This variation might suggest that the genus Singhiella is currently unsatisfactorily defined.
Table 2. Characteristics of Singhiella species from Taiwan.
Species Colour Thoracic Tilelike Pairs of s.m. setae Tips of Posteriormost
tracheal structures (excluding cau submarginal s.m. setae
pores dal setae) setae
6. S. piperis yellow to brown present present 10 spiny submargin 7. S. subrotunda yellowish absent present 12 spiny submargin
but small
8. S. tetrastigmae yellowish present present 15 capitate caudal ridges 9. S. vanieriae pale present present 12 capitate submargin The classification of whiteflies is based primarily on an immature stage, the puparium ( Gill 1990), and adults are usually considered to be morphologically uniform ( BinkMoenen 1991). However, there are several examples of adult characters differing between species, and thus being useful for identification. David & Thenmozhi (1995) provided a comparison table of the clasper setation for four Lipaleyrodes species. Guimarães (1996) described lateral ovipositor and cement gland structure of seven species ( Aleurothrixus floccosus (Maskell) , Aleyrodes proletella Linnaeus , Bemisia tabaci (Gennadius) , Dialeurodes citri (Ashmead) , Parabemisia myricae (Kuwana) , Siphoninus phillyreae (Haliday) and Trialeurodes vaporariorum (Westwood)) but omitted other useful adult morphological characters. Similarly, Calvert et al. (2001) described the length ratios of antennal segments and the position of antennal sensoria for five species belonging to Bemisia , Rhachisphora and Aleurotrachelus . Moreover, the generic definitions of Dialeurodes and Massilieurodes are well supported by the cladistic analysis of Jensen (2001), but Singhiella is much less well supported. Our observations on adult morphology, such as the antennal sensoria and male genitalia, support the genera Dialeurodes and Massilieurodes , but the diversity among the species assigned to Singhiella does not support the suggestion that they belong to the same genus.
After extensive study of whitefly specimens, we have found additional adult characters that could be used to define species. For example, the three related Dialeurodes species, D. agalmae , D. citri and D. daphniphylli , have very similar puparia but the adults differ in the numbers of antennal sensorial plaques, the numbers of ommatidia between upper and lower halves of the compound eyes, the shapes of abdominal wax plates, also the vasiform orifice, claspers, cement glands and lateral view of aedeagus, even the chaetotaxy of male and female genitalia and the chaetotaxy of mesotibia and metatibia. More importance needs to be placed on these adult character states in considering the systematic relationships between whitefly species than has been done hitherto, and adult morphological characteristics need to be taken into consideration in studies on whitefly taxonomy.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |