
Miniopterus egeri, Goodman, Steven M., Ramasindrazana, Beza, Maminirina, Claudette P., Schoeman, Corrie & Appleton, Belinda, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.202738 |
|
DOI |
https://doi.org/10.5281/zenodo.3860372 |
|
persistent identifier |
https://treatment.plazi.org/id/762D87F2-4F5E-FFA6-FF40-D384FD05FB26 |
|
treatment provided by |
Plazi |
|
scientific name |
Miniopterus egeri |
| status |
sp. nov. |
Miniopterus egeri View in CoL sp. nov.
Eger’s long-fingered bat Minioptère d’Eger
( Figures 4–7 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Synonyms
Miniopterus manavi Peterson, Eger & Mitchell, 1995 View in CoL , in part
Miniopterus petersoni Goodman, Bradman, Maminirina, Ryan, Christidis & Appleton, 2008 View in CoL , in part Miniopterus cf. petersoni Goodman, Bradman, Maminirina, Ryan, Christidis & Appleton, 2008 View in CoL , in part
Holotype. FMNH 209160, adult female in reproductive condition, lactating and with one placental scar, body preserved in 12% formalin and subsequently transferred to 70% ethanol, skull removed and cleaned by Dermestidae beetles. The skull is in fine condition. Original field number Steven M. Goodman ( SMG) 16602. This specimen was used in the morphological and molecular analyses, and bioacoustic information is also available.
Referred specimens (specimens used in molecular analysis are marked in bold). Province de Fianarantsoa: USNM 448938 -448939, 0.5 km N Kianjavato, 21º23'S, 47º52'E, 300 m, 12 August 1987, G.K. Creighton and L.H. Emmons; FMNH 187663-187664, Kianjavato, FOFIFA Station, 21º22’674”S, 47º52’152”E, 75 m, 25 September 2005, S.M. Goodman and F. Ratrimomanarivo; USNM 449204-449205, 2 km NE Andrambovato, 21º30'S, 47º54'E, 550 m, 19 August 1988, G.K. Creighton and J.M. Ryan; USNM 449202 -449203, Vatovavy, 9 km ESE Kianjavato, 21º24'S, 47º57'E, 150 m, 30 July 1988, G.K. Creighton and J.M. Ryan. Province de Toamasina: FMNH 202474, Forêt de Sahafina, 9.5 km W Brickaville, 18º48.442’S, 48º58.924’E, 105 m, 9 December 2007, S.M. Goodman, M. Ruedi, and N. Weyeneth; FMNH 209159, 209160, 209161, 209168, Forêt de Sahafina, 9.5 km W Brickaville, 18º48’37”S, 48º58’48”E, 50 m, 5 & 10 December 2009, S.M. Goodman and B. Ramasindrazana; FMNH 202519, 202520, 202521, Poste Forestier de Farankaraina, 7.5 km SW Mahalevona, 15º25.694’S, 49º50.717’E, 50 m, 24 November 2007, M. Ruedi and S.M. Goodman; FMNH 202522, 1.1 km S Ambanizana, along Androka River, 15º38.053’S, 49º58.114’E, 5 m, 25 November 2007, S.M. Goodman, M. Ruedi, and N. Weyeneth. Some of these specimens were previously used in the diagnosis of M. petersoni (Goodman et al. 2008) .
Etymology. This species in named in honor of Dr. Judith Eger, Senior Curator, Department of Mammalogy, Royal Ontario Museum, Toronto, for her contribution to taxonomic studies of Old World bats, including Madagascar (see Monadjem et al. 2010 for further details).
Diagnosis. A moderately small species of Miniopterus with mixed medium and dark brown colored pelage ( Figure 4 View FIGURE 4 ). The holotype has a forearm length of 39 mm and in the species this measurement ranges from 37–40 mm (mean= 38.5 mm). Tragus relatively short, 5–6 mm in length, slightly constricted on lower distal side, and distal portion notably thickened ( Figure 5 View FIGURE 5 ). From dorsal view, cranial palatal emargination with angularly tapered base, forming a slightly open “V-shaped” ( Figure 6 View FIGURE 6 ). Rostrum and interorbital region narrow. Rostral depression narrow, relatively shallow and long. Notable medial inflation of supraorbital ridge. Based on molecular characters, M. egeri forms a distinct clade from other Malagasy moderately small members of the genus.
Description. We have directly compared the holotypes of M. egeri (FMNH 209160) and M. petersoni (FMNH 194136) for external and cranio-dental characters, as well as series referable to these two species and other moderately small Malagasy members of the genus, including M. sororculus and M. majori .
External characters. A moderately small Miniopterus with a tail less than 50% of total length ( Table 5 View TABLE 5 ). In the holotype, which was preserved in fluid in late 2009, the dorsal and ventral pelage is slightly long, dense and a mix of medium brown interspersed with a distinctly lighter brown colored fur ( Figure 4 View FIGURE 4 ). This individual was compared to five specimens prepared as dry skins (USNM 448938-448939, 449203-449205). USNM 448939, in which the back and upper ventrum coloration approaches a dark brown, and USNM 449204, in which the back is a slightly lighter brown, show slightly different fur coloration than the holotype. Across the type series, the wing membrane and uropatagium are dark brownish-black, largely naked, and show no noticeable change in coloration across their surface area.
The ear average length of the holotype of M. egeri is 10 mm, and the range for the ten specimens referred to this species is 10–11 mm (average 10.6 mm), which is smaller than the average values of 11.7 mm for animals referred to M. petersoni ; these differences are statistically significant ( Table 5 View TABLE 5 ; all measured by same field collector). The tragus length in the holotype of M. egeri is 6 mm and this species shows little variation with all animals falling with the range 5–6 mm (average 5.9 mm); it does not show extensive overlap with M. petersoni with minimum–maximum values of 6–7 mm (average 6.7 mm; this difference is statistically significant ( Table 5 View TABLE 5 ; all measured by same field collector).
The tragus of M. egeri is notably thick along the shaft, slightly constricted on lower distal side, the length of the proximal edge has a distinct flange that folds slightly anterior-medially, and the downward deflected distal tip is notably thickened and slightly fleshy ( Figure 5 View FIGURE 5 a). In comparison, M. petersoni has a distinctly thinner tragus shaft, the constriction of the lower proximal portion notably more pronounced, and the distal tip has a slightly raised and rounded structure ( Figure 5 View FIGURE 5 b). The tragus in M. sororculus is similar in length to M. petersoni ( Table 5 View TABLE 5 ), but distinctly more spatulate-shaped and without a notable tapered distal head ( Figure 5 View FIGURE 5 c).
On the basis of most external measurements, M. egeri is more diminutive than other moderately small Malagasy Miniopterus ( Table 5 View TABLE 5 ). For example, the average forearm length in M. egeri is 38.5 mm (range 37–40 mm), M. petersoni 40.0 mm (range 39–41 mm), M. sororculus 43.5 mm (range 42–45 mm), and M. majori 45.5 mm (range 44–47 mm). The difference between the first two taxa is statistically significant. In contrast, the hindfoot length in M. egeri and M. petersoni broadly overlap, but this measurement is slightly longer in M. sororculus and notably longer in M. majori ( Table 5 View TABLE 5 ).
Cranio-dental characters. The skull of M. egeri is notably short with a greatest skull length of 14.3 mm on average (range 13.9–14.8 mm), overlapping with M. petersoni with its average length of 14.5 mm (range 14.1–14.9 mm); this slight difference is statistically significant ( Table 6 View TABLE 6 ). Miniopterus sororculus has approximately the same skull length as M. egeri and M. petersoni , but M. majori is notably longer. Miniopterus egeri has a notably short and narrow rostrum and not a particularly bulbous braincase, similar to M. petersoni ( Figure 6 View FIGURE 6 ), although there are a few statistical differences between these two taxa ( Table 6 View TABLE 6 ). In M. egeri , the interorbital region is narrow and the rostral depression tends to be proportionately narrow and relatively shallow across most of its length as compared to M. petersoni . In the former species, the supraorbital ridge shows a notable medial inflation, similar to M. petersoni ( Figure 6 View FIGURE 6 ). When viewed from above, the palatal emargination in M. egeri is relatively narrow and forms a slightly open “V-shape”; in contrast, the emargination in M. petersoni is a more closed “V-shape” ( Figure 6 View FIGURE 6 ).
The upper toothrows in M. egeri are relatively short for an animal of its size, largely in parallel, although slightly posteriorly diverging ( Figure 7 View FIGURE 7 ); the upper toothrow is similar in length to M. petersoni and M. sororculus ( Table 7 View TABLE 7 ). The lower tooth row in M. egeri is 6.6 mm on average (range 6.4–7.0 mm), which is shorter than the average of M. petersoni at 6.8 mm (range 6.4–7.0), and this difference is statistically significant ( Table 7 View TABLE 7 ). Dental formula I 2 /3 C 1/1 P 2/3 M 3/3, comprising the adult dentition of 36 teeth. First upper premolar (P2) is relatively small and with more simple cusp morphology than second upper premolar (P3), typical of members of this genus ( Koopman 1994). The dentition of M. egeri is similar to M. petersoni and other moderately small Miniopterus .
Distribution and conservation status. Over the past decade, largely based on faunal surveys and associated specimens and tissues obtained, important advances have been made in understanding the species limits and distribution of Malagasy region Miniopterus . On the basis of current information, which by no means should be considered complete, it is possible to delineate the distributions of members of this genus occurring in the more mesic eastern portions of the island ( Goodman 2011): M. brachytragos Goodman, Maminirina, Bradman, Christidis, Appleton, 2009 —largely a western species but has been captured near Maroantsetra from near sea level; M. gleni —Masoala Peninsula to Tolagnaro and scattered records from the Central Highlands from sea level to 1200 m; M. egeri— Masoala Peninsula to Vatovavy from 50 to 550 m; M. majori —largely confined to the Central Highlands, but has also been found near Maroantsetra south to Tolagnaro from sea level to over 1550 m; M. manavi— Central Highlands, near Fandriana, south to Ranomafana (Ifanadiana) and Vinantelo from 900 to 1500 m; M. peter-soni— lowland portions of the southeast in the vicinity of Tolagnaro from 10 to 550 m; and M. sororculus— Central Highlands from 950 to 2200 m. Hence, amongst these small to medium sized species, with the exception of M. sororculus and M. majori , none are known to occur in sympatry in the east. Miniopterus egeri is allopatric with M. petersoni , which in turn do not have overlapping elevational distributions with M. manavi . Approximately 300 km separate the known northern limit of M. petersoni and the southern limit of M. egeri ( Figure 1 View FIGURE 1 ). Medium to small Miniopterus are known form this intermediate zone (e.g., Midongy-Sud), but based on molecular genetics data represent other clades than those species mentioned herein (Appleton et al. unpublished data).
Most of the sites where M. egeri occurs are within intact or partially degraded forest ecosystems and few details are available about this species’ natural history. On the basis of current information, it is known from scattered sites occurring across two-thirds of Madagascar’s lowland area from near sea level to 550 m, often associated with disturbed forests with mixed native and introduced trees or at the ecotone between degraded anthropogenic habitats and native forest. A day roost site of this species was found in a natural rock shelter in the Poste Forestier de Farankaraina surrounded by slightly disturbed natural lowland humid forest. At least in the short-term, this species does not appear to face a risk of dramatic population decline. However, further information is needed on its distribution and size of existing colonies to properly assess its conservation status.
T–statistics for
M. egeri compared T=2.75 to M. petersoni n.s. n.s. n.s. n.s. P=0.009 Across its range from the Maroantsetra region south to near Vatovavy ( Figure 1 View FIGURE 1 ), a zone of approximately 700 km in length, there is no noticeable variation in the morphometrics of M. egeri . In Tables 5 View TABLE 5 , 6 View TABLE 6 , and 7, different external, cranial, and dental measurements (respectively) of specimens are segregated into two different geographic zones, the southeast (Vatovavy to Sahafina) and northeast (Masoala Peninsula). In general, although sample sizes are not large, measurements are consistent between the two zones. Further sampling in other areas, specifically the zone north of Toamasina and south of Maroantsetra, will need to be conducted to verify if M. egeri has a continuous distribution across much of the eastern portion of the island.
In their description of M. petersoni, Goodman et al. (2008) presented several characters that clearly separated that species from the other Afro-Malagasy Miniopterus . Subsequently, a number of species have been described or elevated from subspecies level known from Madagascar and surrounding islands, which are distinctly larger ( M. griffithsi ; Goodman et al. 2010) or smaller ( M. aelleni Goodman, Maminirina, Weyeneth, Bradman, Christidis, Ruedi & Appleton, 2009 , M. brachytragos , and M. mahafaliensis Goodman, Maminirina, Bradman, Christidis, Appleton, 2009 ) than M. petersoni and its morphologically similar species M. egeri . All of these recently recognized Miniopterus species can be differentiated from M. egeri based on tragus shape and other morphological characters and, hence, these taxa do not need to be considered further. The remaining needed comparison is the morphological distinction between M. petersoni and M. egeri .
Goodman et al. (2008) noted that specimens allocated to M. petersoni from the Tolagnaro region, which includes the holotype locality, showed some measurement and morphological differences to those obtained from the central southeast, as well as the northeast portions of the island. As molecular genetics have been of primary importance for the recognition of cryptic Miniopterus species from the Madagascar region (see for example, Goodman et al. 2009a; Weyeneth et al. 2008), and given that tissue samples were not associated with the Goodman et al. (2008) study for specimens from the central southeast and northeast allocated to M. petersoni , the resolution of their specific status was uncertain. On the basis of the comparisons presented above, it is now clear that populations previously assigned to M. petersoni from the zone between the central southeast (Vatovavy to Sahafina) and northeast (Masoala Peninsula) show genetic sequence divergence as well as morphological characters, such as tragus shape, that separate them from extreme southeastern populations of M. petersoni . There also appear to be bioacoustic differences that separate M. egeri and M. petersoni . However, echolocation data recorded in different natural habitats are needed to confirm this.
To further explore morphological differences between genetically or morphologically similar Malagasy Miniopterus spp. we conducted PCAs on external, cranial, and dental measurements of M. egeri , M. petersoni , M. sororculus , M. majori , and the diminutive M. manavi sensu stricto. The comparisons of external, cranial, and dental measurements generally showed clear separation of specimens assigned to M. sororculus , M. majori , M. manavi , and members of the M. egeri / M . petersoni complex; only the cranial analysis is presented here ( Figure 8). The first two unrotated principal components (PCs) accounted for 94% of the total variance of the cranial morphology, and separated the specimens into four groups: M. sororculus , M. majori , M. manavi , and the M. egeri / M . petersoni complex ( Figure 8; Table 8 View TABLE 8 ). All of the eight cranial variables showed heavy loadings on PC1, but only three cranial variables, POB, GBW, and MAND loaded high on PC2. These patterns are interpreted as follows. PC1 was a measure of cranial size and bats that loaded low on PC1 ( M. majori ) had relatively larger skulls than bats that loaded high on PC1 ( M. egeri and M. manavi ). PC2 was a measure of cranial shape, and bats that loaded low on PC2 ( M. sororculus ) had relatively broader brain cases and crania with shorter mandibular lengths compared to bats that loaded high on PC2 ( M. petersoni ). In the case of the sister taxa M. egeri and M. petersoni , which have allopatric distributions, there is notable overlap in the PCA plot of cranial variables for PC1 and PC2. In contrast, the sympatrically occurring and phylogenetically distinct M. majori and M. sororculus ( Figure 2 View FIGURE 2 ) show complete separation in the PCA analysis for both PC1 and PC2. These differences in size and shape might be associated with interspecific competition between Miniopterus spp. of similar ecology and overlapping distributions.
In a more practical sense, in the hand M. egeri can be separated from other moderately small to medium-sized Malagasy members of this genus, particularly M. petersoni and M. sororculus , by tragus shape ( Figure 5 View FIGURE 5 ). Based on current information there is no evidence that M. egeri and M. petersoni occur in sympatry. Hence, the geographical zone a given animal was captured is also informative as to its specific designation.
One of the remaining questions is the specific status of the moderately small Miniopterus from the upper slopes of Montagne d’Ambre in the far north that were allocated to M. cf. petersoni by Goodman et al. (2008), but showed notable measural differences with typical M. petersoni from the southeast. The animals from Montagne d’Ambre have relatively broader rostrums, approaching that of M. sororculus . In order to resolve this question, further specimens and associated tissue are needed from the Montagne d’Ambre population.
3,0
2,5
2,0
1,5
1,0
0,5
PC 0,0 2
-0,5
-1,0
-1,5
Miniopterus egeri View in CoL -2,0 M. petersoni View in CoL M. sororculus View in CoL M. majori View in CoL
-2,5 M. manavi View in CoL s. s. -2,0 -1,5 -1,0 -0,5 0,0 0,5 1,0 1,5 2,0 PC 1
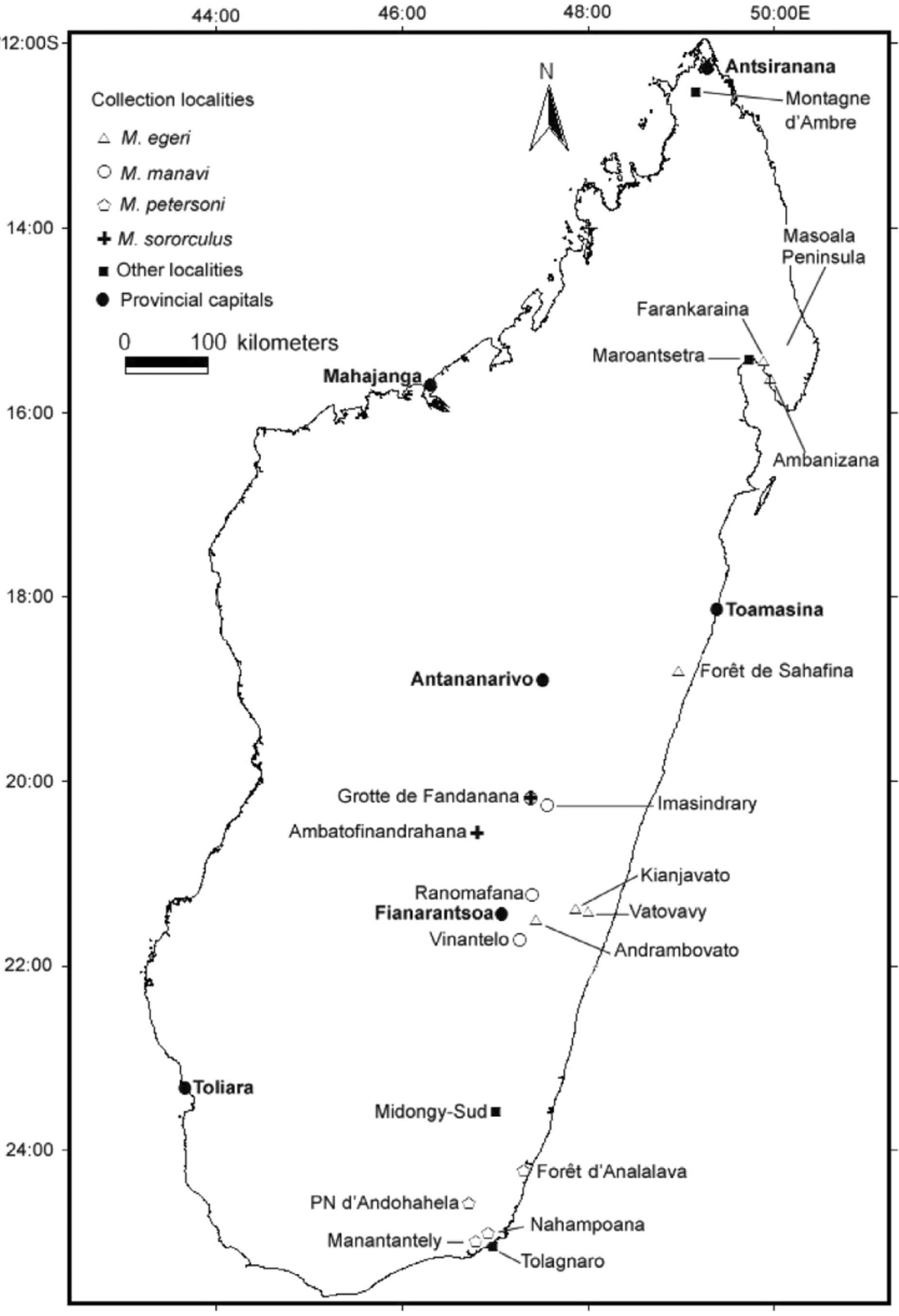
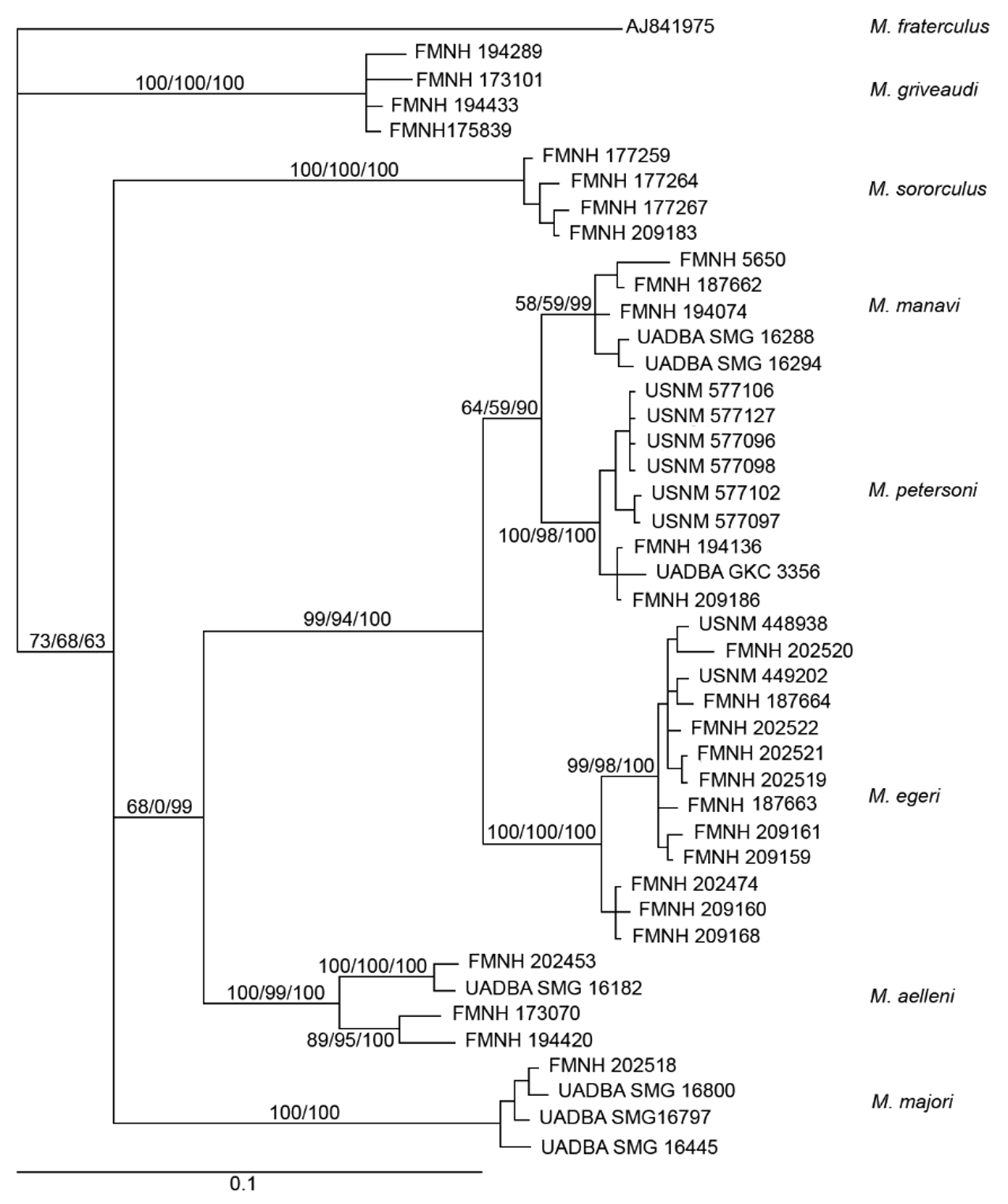

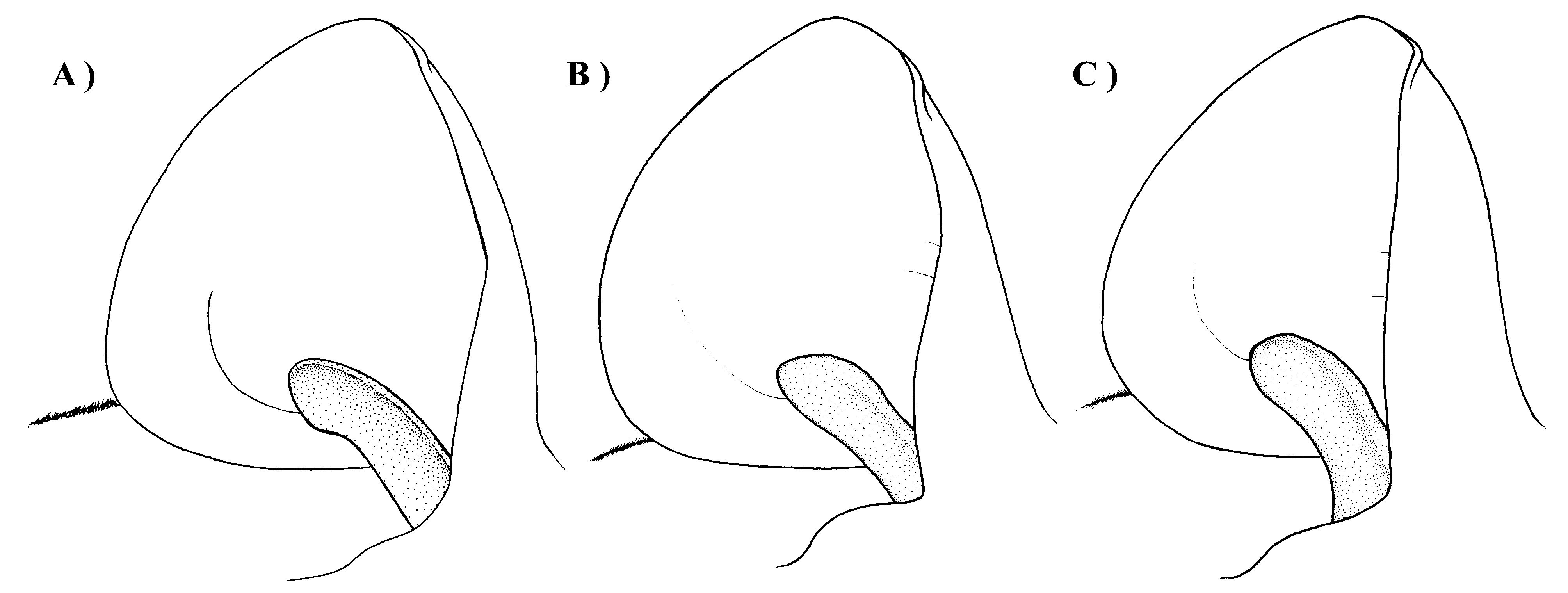
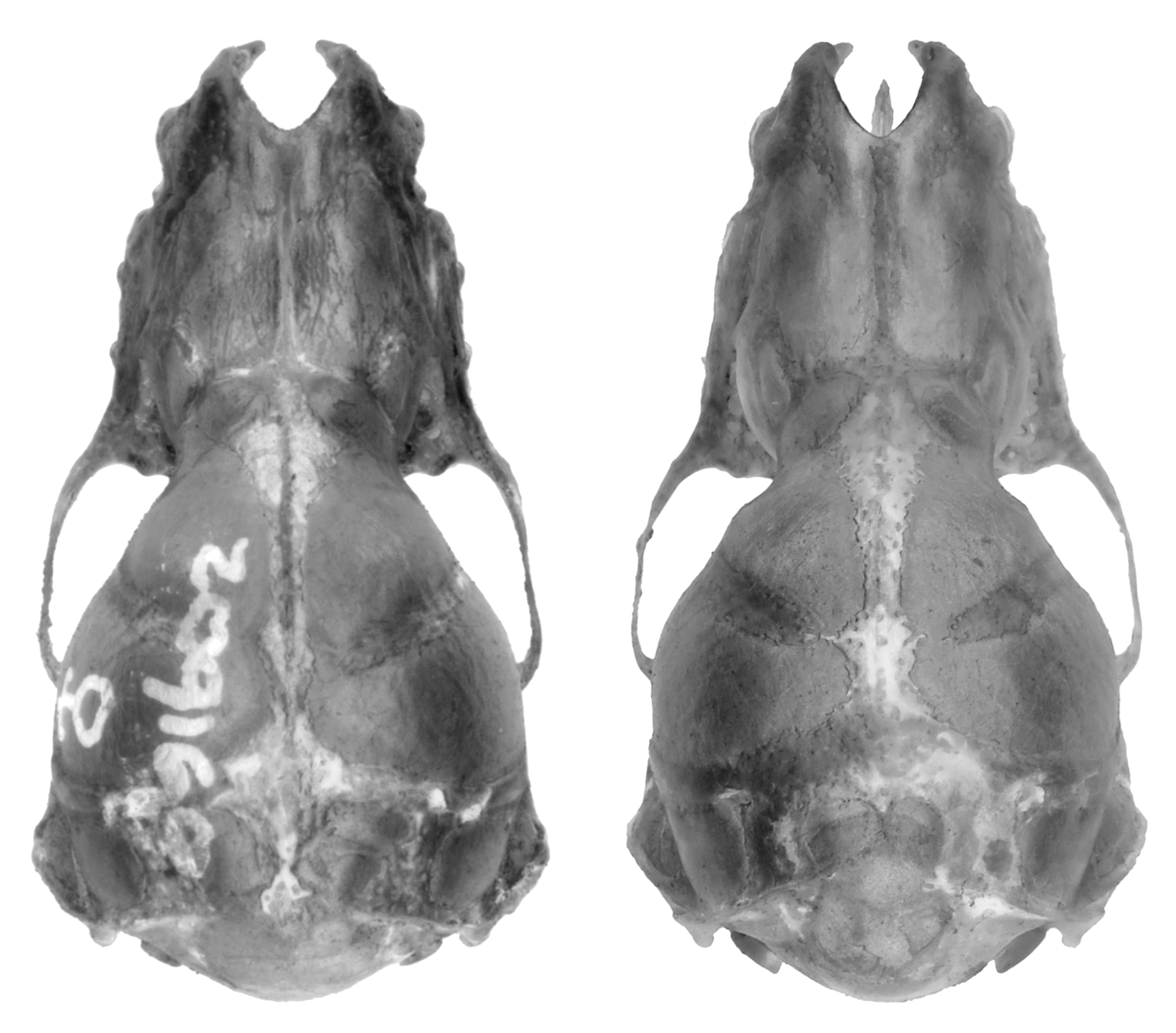
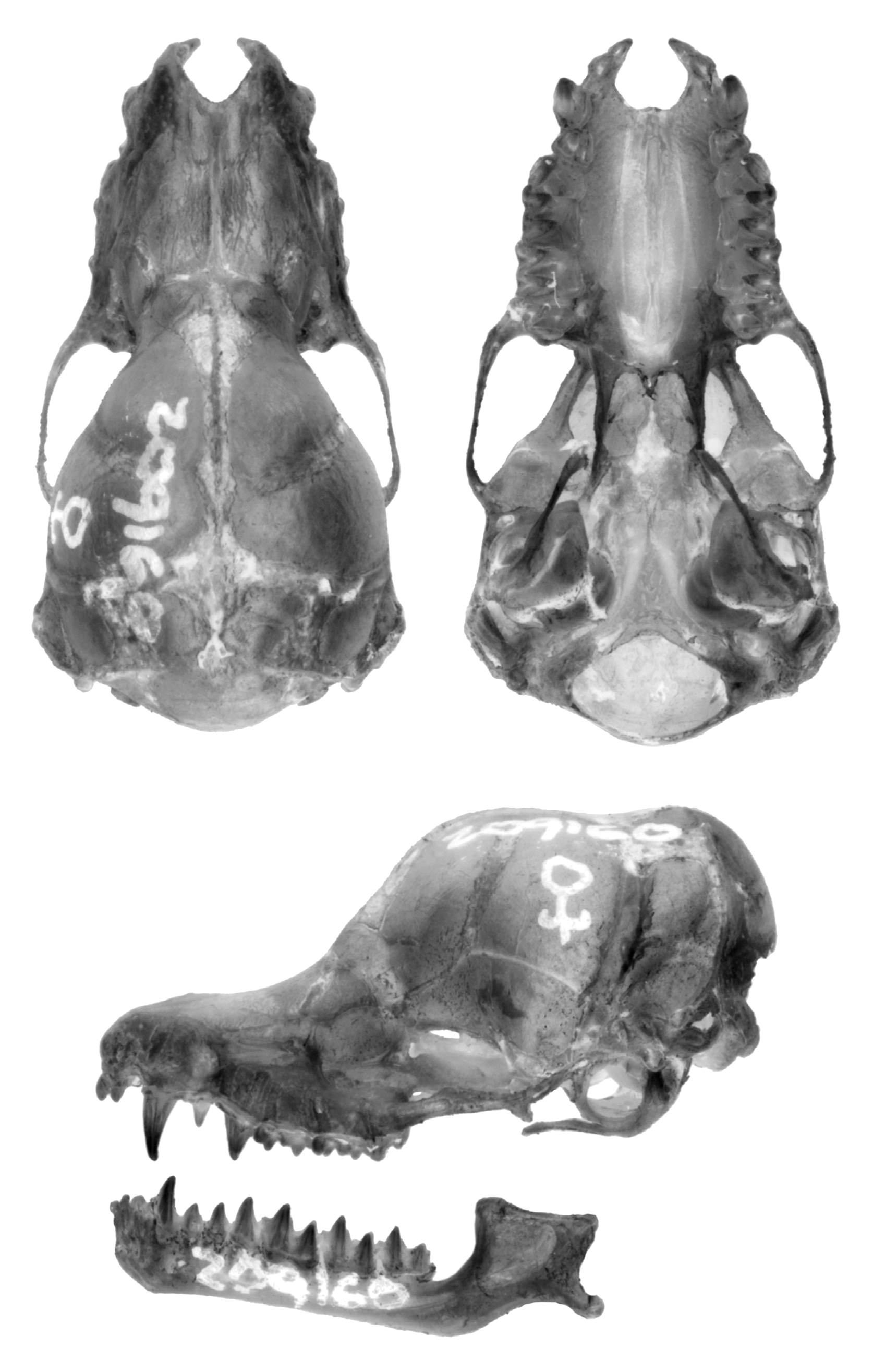
TABLE 5. External measurements (in millimeters) and mass (in grams) of Miniopterus egeri sp. nov., M. petersoni, M. sororculus (Central Highlands), and M. majori. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. Figures in bold are of specimens collected and measured by SMG and those in standard script by a variety of different field collectors. Statistical differences between M. egeri and M. petersoni are examined based on student t-tests, n. s. = not significant.
| Total length | Tail length | Hindfoot length | Tragus length | Ear length | Forearm Body length mass | |
|---|---|---|---|---|---|---|
| M. egeri Holotype FMNH 209160 Ƥ | 91 | 43 | 7 | 6 | 10 | 39 6.1 |
| M. egeri Central east | 92.3 ± 1.51 91–95, n=6 | 41.7 ± 1.51 40–44, n=6 | 6.3 ± 0.82 5–7, n=6 | 5.8 ± 0.41 5–6, n=6 | 10.3 ± 0.52 10–11, n=6 | 38.3 ± 0.76 6.3 ± 0.69 37–39, n=6 5.6–7.6, n=6 |
| M. egeri Northeast | 91.0 ± 2.16 89–94, n=4 | 41.8 ± 0.96 41–43, n=4 | 6.8 ± 0.50 6–7, n=4 | 6.0 ± 0.0 6–6, n=4 | 11.0 ± 0.0 11–11, n=4 | 39.0 ± 0.82 5.4 ± 0.25 38–40, n=4 5.2–5.7, n=4 |
| M. egeri Across range | 91.8 ± 1.81 89–95, n=10 | 41.7 ± 1.25 40–44, n=10 | 6.5 ± 0.71 5–7, n=10 | 5.9 ± 0.32 5–6, n=10 | 10.6 ± 0.52 10–11, n=10 | 38.5 ± 0.73 5.7 ± 0.75 37–40, n=15 4.2–7.6, n=15 |
| M. petersoni Holotype FMNH 194136 3 | 98 | 45 | 6.5 | 6 | 10 | 40.5 6.5 |
| M. petersoni Southeastern Madagascar | 93.9 ± 3.06 89–99, n=15 | 43.6 ± 2.44 39–50, n=15 | 6.0–6.5, n=2 | 6.7 ± 0.46 6–7, n=15 | 11.7 ± 0.82 10–13, n=15 | 40.0 ± 0.70 6.3 ± 0.80 39–41, n=14 5.5–8.2, n=15 |
| T-statistics for M. egeri compared to M. petersoni | n.s. | T=2.06 P =0.05 | n.s. | T=5.19 P <0.00001 | T=3.98 P =0.0006 | T=5.63 P <0.00001 n.s. |
| M. sororculus M. majori | 110.8 ± 2.71 105–115, n=23 115.6 ± 2.42 112–120, n=39 | 54.7 ± 1.84 51–58, n=23 55.5 ± 1.90 51–60, n=39 | 6.7 ± 0.65 6–8, n=23 7.7 ± 0.54 7–9, n=37 | 6.7 ± 0.65 6–8, n=23 7.3 ± 0.46 7–8, n=39 | 10.7 ± 0.62 10–12, n=23 11.9 ± 0.47 11–13, n=39 | 43.5 ± 0.85 7.9 ± 0.62 42–45, 7.0–9.1, n=23 n=22 45.5 ± 0.82 9.7 ± 0.93 44–47, 8.4–12.5, n=39 n=37 |
TABLE 6. Cranial measurements (in millimeters) of adult Miniopterus egeri sp. nov., M. petersoni, M. sororculus (Central Highlands), and M. majori. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. See Methods and materials for an explanation of variable acronyms. Statistical differences between M. egeri and M. petersoni are examined based on student t-tests, n. s. = not significant.
| GSKL | CIL | ZYGO | POB MAST GBW | PAL | LW | MAND |
|---|---|---|---|---|---|---|
| M. egeri Holotype FMNH 209160 Ƥ 14.8 | 14.3 | 7.7 | 3.3 7.9 7.2 | 5.8 | 4.7 | 10.4 |
| M. egeri 14.4 ± 0.38 Central east 13.9–14.8, n=9 | 13.7 ± 0.38 13.3–14.3, n=8 | 7.5 ± 0.15 7.3–7.7, n=7 | 3.1 ± 0.13 7.6 ± 0.23 6.9 ± 0.20 2.9–3.3, 7.3–7.9, 6.7–7.2, n=9 n=9 n=9 | 5.4 ± 0.31 5.0–5.9, n=9 | 4.5 ± 0.16 4.2–4.7, n=10 | 10.2 ± 0.38 9.7–11.0, n=10 |
| M. egeri 14.2 ± 0.18 Northeast 14.0–14.4, n=4 | 13.5 ± 0.12 13.4–13.6, n=4 | 7.7 ± 0.06 7.6–7.7, n=4 | 3.3 ± 0.06 7.6 ± 0.08 7.0 ± 0.05 3.2–3.3, 7.5–7.7, 6.9–7.0, n=4 n=4 n=4 | 5.1 ± 0.06 5.0–5.2, n=3 | 4.6 ± 0.10 4.5–4.7, n=4 | 9.9 ± 0.10 9.8–10.0, n=4 |
| M. egeri 14.3 ± 0.30 Across range 13.9–14.8, n=13 | 13.6 ± 0.32 13.3–14.3, n=12 | 7.6 ± 0.13 7.3–7.7, n=11 | 3.2 ± 0.15 7.6 ± 0.20 6.9 ± 0.17 2.9–3.3, 7.3–7.9, 6.7–7.2, n=13 n=13 n=13 | 5.3 ± 0.31 5.0–5.9, n=12 | 4.5 ± 0.15 4.2–4.7, n=14 | 10.1 ± 0.34 9.7–11.0, n=14 |
| M. petersoni Holotype FMNH 194136 3 14.7 | 14.1 | 7.8 | 3.2 7.8 7.0 | 5.6 | 4.6 | 10.3 |
| M. petersoni 14.5 ± 0.19 Southeastern 14.1–14.9, Madagascar n=34 | 13.9 ± 0.22 13.2–14.1, n=34 | 7.7 ± 0.16 7.4–8.1, n=28 | 3.1 ± 0.12 7.6 ± 0.15 6.8 ± 0.19 2.9–3.4, 7.3–7.9, 6.5–7.3, n=34 n=31 n=31 | 5.4 ± 0.12 5.2–5.6, n=31 | 4.5 ± 0.17 4.2–4.8, n=33 | 10.2 ± 0.20 9.8–10.6, n=31 |
| T-statistics for M. egeri compared to T=2.33 M. petersoni P=0.03 | T=2.76 P=0.008 | n.s. | n.s. n.s. n.s. | n.s. | n.s. | n.s. |
| M. sororculus 14.5 ± 0.24 13.8–14.9, n=52 | 14.0 ± 0.25 13.5–14.4, n=52 | 8.1 ± 0.15 7.7–8.3, n=44 | 3.4 ± 0.11 8.0 ± 0.19 7.4 ± 0.14 3.1–3.6, 7.7–8.2, 7.1–7.7, n=54 n=52 n=51 | 5.4 ± 0.20 5.2–6.0, n=41 | 4.7 ± 0.16 4.5–4.9, n=55 | 10.0 ± 0.15 9.7–10.3, n=28 |
| M. majori 15.4 ± 0.26 14.9–15.8, n=40 | 15.1 ± 0.19 14.7–15.4, n=40 | 8.5 ± 0.14 8.2–8.8, n=40 | 3.6 ± 0.11 8.3 ± 0.16 7.7 ± 0.14 3.3–3.8, 7.9–8.7, 7.4–8.0, n=40 n=40 n=40 | 5.9 ± 0.12 5.7–6.3, n=39 | 5.3 ± 0.16 4.9–5.5, n=40 | 10.9 ± 0.20 10.5–11.3, n=36 |
TABLE 7. Dental measurements (in millimeters) of Miniopterus egeri sp. nov., M. petersoni, M. sororculus (Central Highlands), and M. majori. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. See Methods and materials for an explanation of variable acronyms. Statistical differences between M. egeri and M. petersoni are examined based on student t-tests, n. s. = not significant.
| M. egeri Holotype FMNH 209160 Ƥ | I1-M3 6.8 | C-M3 5.6 | C1-C1 4.0 | M3-M3 5.8 | i1-m3 7.0 |
|---|---|---|---|---|---|
| M. egeri Central east | 6.4 ± 0.23 6.2–6.8, n=10 | 5.4 ± 0.14 5.2–5.6, n=10 | 3.9 ± 0.11 3.8–4.1, n=10 | 5.5 ± 0.22 5.2–5.8, n=10 | 6.7 ± 0.21 6.4–7.0, n=10 |
| M. egeri Northeast | 6.4 ± 0.10 6.3–6.5, n=4 | 5.4 ±0.13 5.2–5.5, n=4 | 4.0 ± 0.08 3.9–4.1, n=4 | 5.5 ± 0.06 5.4–5.5, n=4 | 6.5 ± 0.03 6.5–6.6, n=4 |
| M. egeri Across range | 6.4 ± 0.20 6.2–6.8, n=14 | 5.4 ± 0.13 5.2–5.6, n=14 | 4.0 ± 0.10 3.8–4.1, n=14 | 5.5 ± 0.18 5.2–5.8, n=14 | 6.6 ± 0.19 6.4 –7.0, n=14 |
| M. petersoni Holotype FMNH 194136 3 | 6.7 | 5.6 | 3.9 | 5.8 | 6.8 |
| M. petersoni Southeastern Madagascar | 6.5 ± 0.14 6.1–6.7, n=34 | 5.4 ± 0.13 5.1–5.6, n=34 | 3.9 ± 0.11 3.5–4.1, n=34 | 5.5 ± 0.13 5.1–5.8, n=34 | 6.8 ± 0.12 6.4–7.0, n=34 |
TABLE 8. Factor loadings from principal component analysis of log-transformed cranial measurements of specimens of Miniopterus egeri sp. nov., M. manavi sensu stricto, M. petersoni, M. sororculus, and M. majori. A graphical representation of the first two factors is presented in Figure 8. See Methods and materials for the acronym definitions.
| Variable | PC 1 | PC 2 | PC 3 |
|---|---|---|---|
| GSKL | -0.934 | 0.269 | -0.159 |
| CIL | -0.962 | 0.204 | -0.106 |
| ZYGO | -0.971 | -0.132 | -0.051 |
| POB | -0.884 | -0.376 | 0.106 |
| MAST | -0.967 | -0.160 | -0.073 |
| GBW | -0.918 | -0.342 | -0.087 |
| LW | -0.905 | 0.138 | 0.387 |
| MAND | -0.893 | 0.401 | 0.007 |
| Eigenvalue | 6.917 | 0.595 | 0.213 |
| Proportion of total explained variation | 86.4% | 93.8% | 96.5% |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Miniopterus egeri
| Goodman, Steven M., Ramasindrazana, Beza, Maminirina, Claudette P., Schoeman, Corrie & Appleton, Belinda 2011 |
Miniopterus petersoni
| Goodman, Bradman, Maminirina, Ryan, Christidis & Appleton 2008 |
Miniopterus cf. petersoni
| Goodman, Bradman, Maminirina, Ryan, Christidis & Appleton 2008 |
Miniopterus manavi
| Peterson, Eger & Mitchell 1995 |