
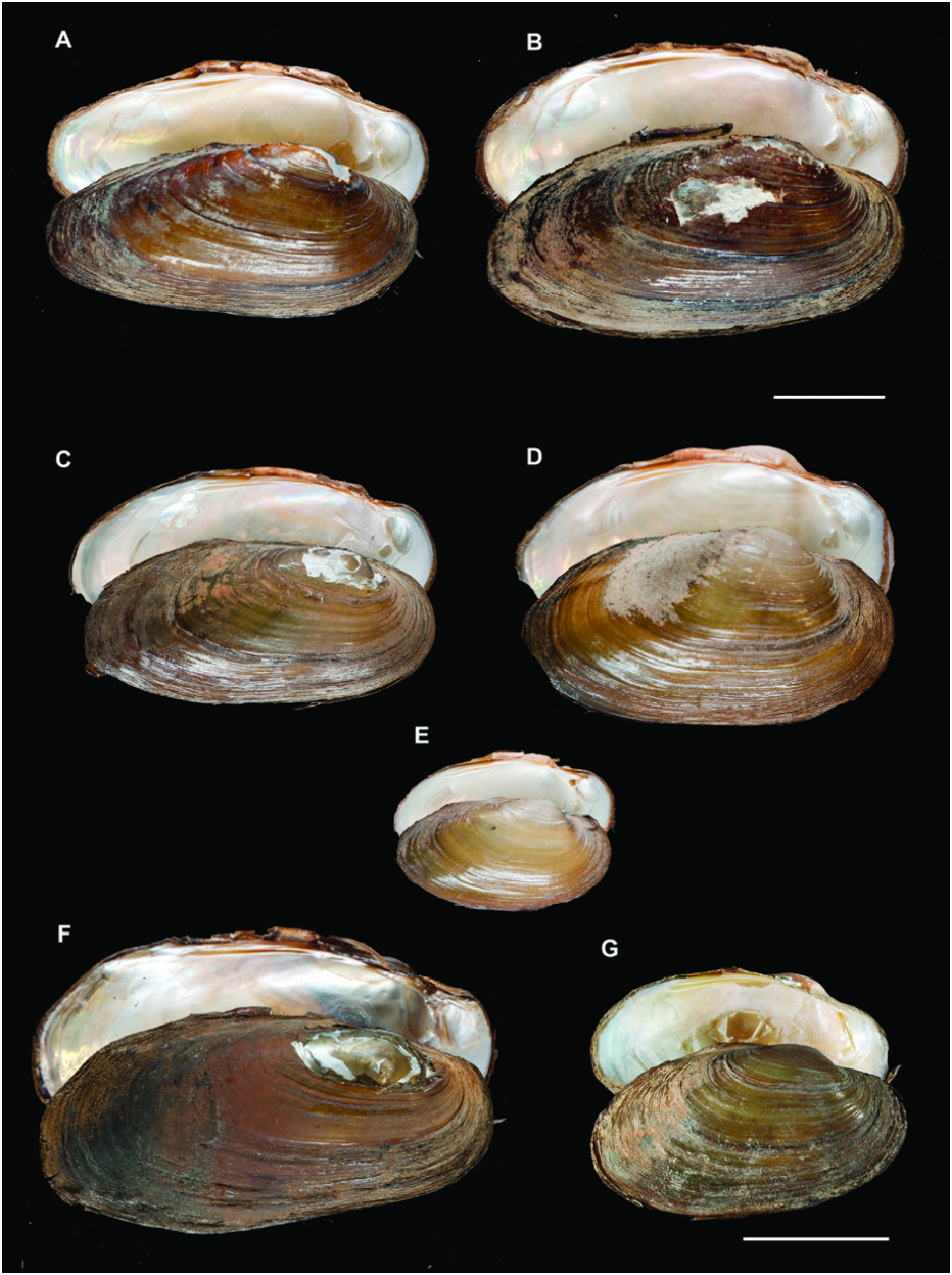
Unio foucauldianus, Pallary, 1936
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlx039 |
|
DOI |
https://doi.org/10.5281/zenodo.5713995 |
|
persistent identifier |
https://treatment.plazi.org/id/744587AC-FFC2-D520-9D2C-51ADFBFE0735 |
|
treatment provided by |
Carolina |
|
scientific name |
Unio foucauldianus |
| status |
|
The ( U. foucauldianus + U. delphinus) + U. tigridis, U. ravoisieri, U. mancus, U. elongatulus, U. pictorum lineage
This lineage was the most complex within the genus ( Fig. 1 View Figure 1 ). All morphospecies were recovered as monophyletic groups, although some had high levels of within-clade diversity and phylogenetic structuring. The relationships among lineages, however, were not well supported.
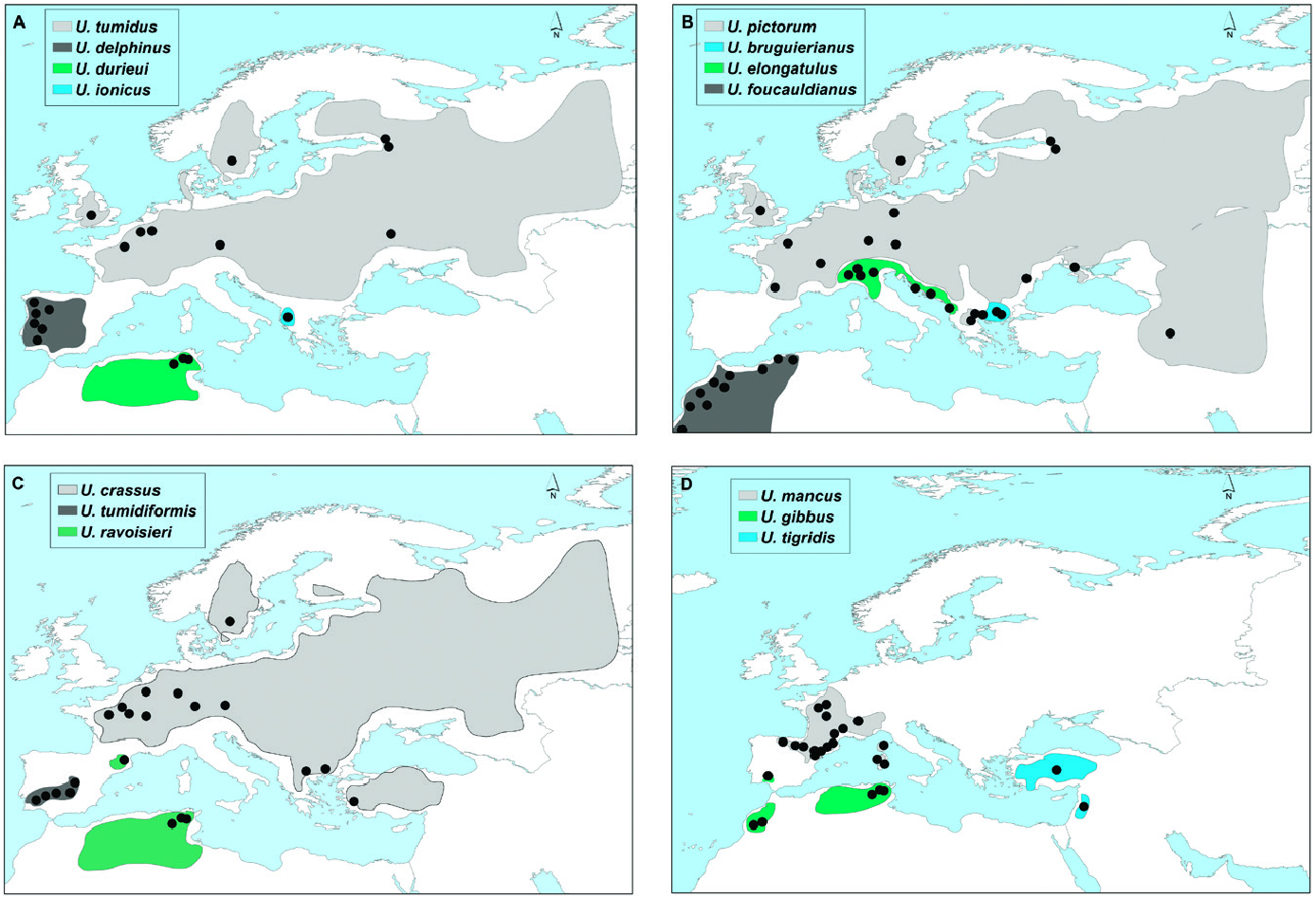
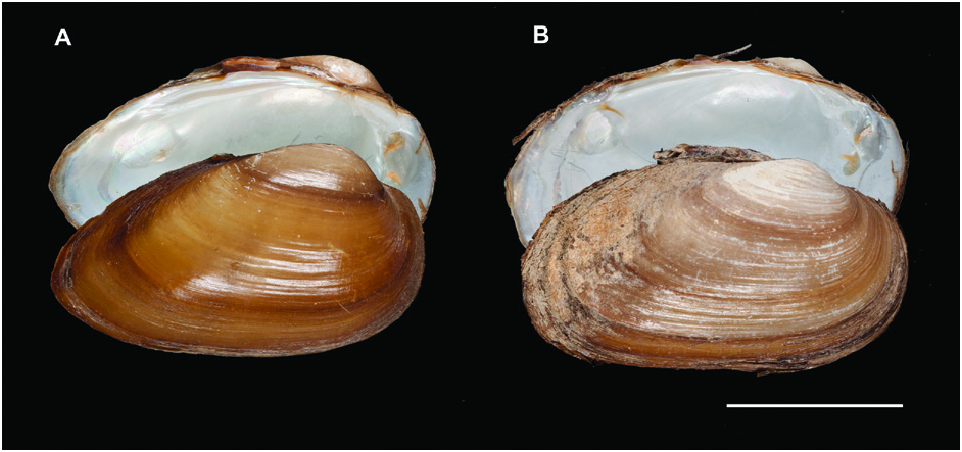
The first well-supported clade of this lineage included two closely related morphospecies with a genetic distance of 3.32% ( Fig. 1 View Figure 1 ; Table 1 View Table 1 ): U. delphinus from Atlantic Iberia and U. foucauldianus from Atlantic and Mediterranean Morocco ( Fig. 2 View Figure 2 ). The network analysis showed a minimum of 24 steps separating the two species (Supporting Information, Fig. S4 View Figure 4 ). There was greater Atlantic/South differentiation in U. delphinus (no shared haplotypes between the 2 regions and up to 24 substitutions among the most differentiated haplotypes) than in U. foucauldianus (in this case, the main haplotype was found in both the Atlantic and Mediterranean basins).
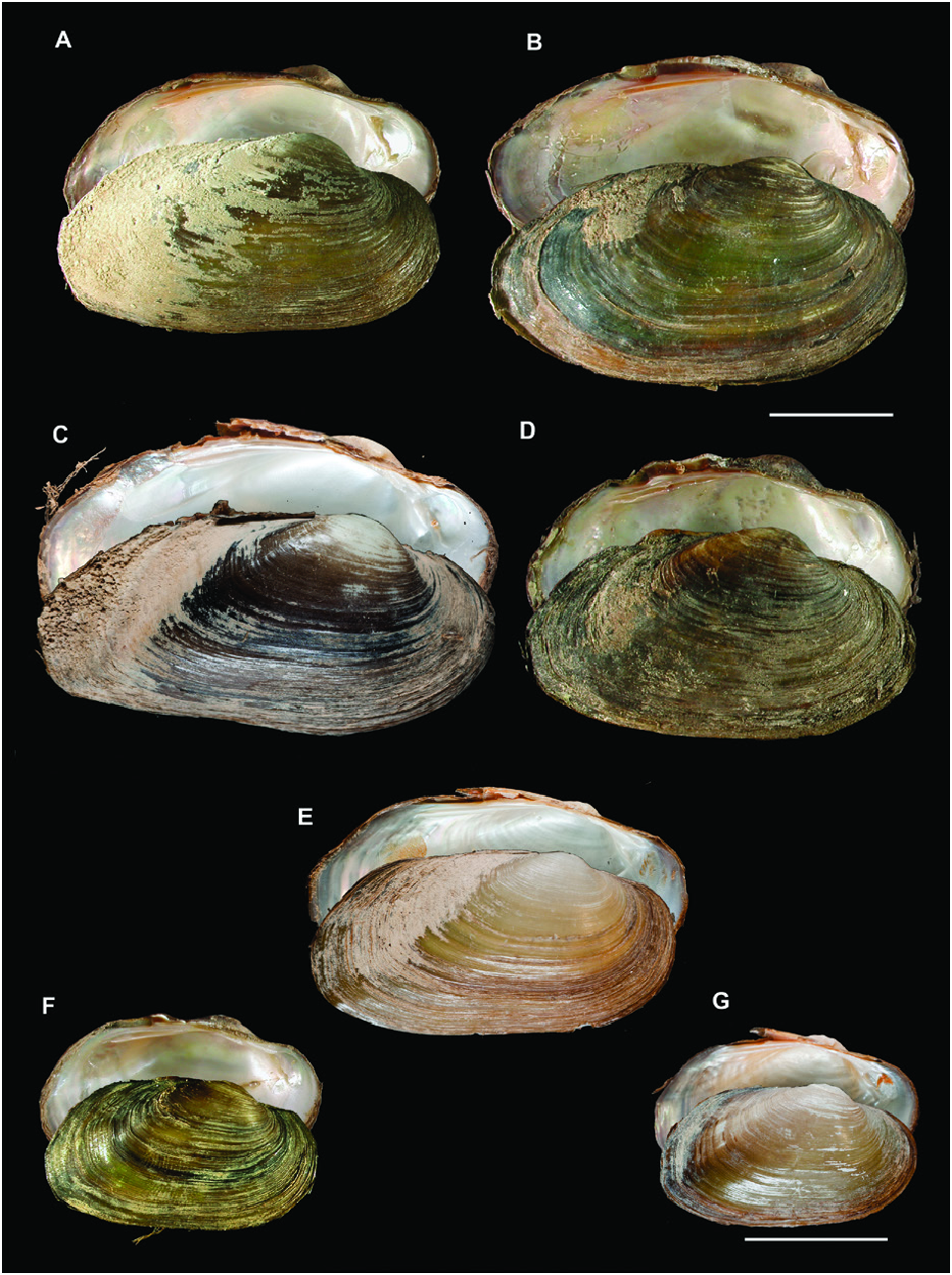
Many of the U. delphinus and all of the U. foucauldianus specimens from Mediterranean Morocco had an elongated shell with a pictorum shape, while specimens from the Atlantic rivers of Morocco and southern Spain were more rounded, resembling the mancus shape ( Figs 4 View Figure 4 , 5 View Figure 5 ).
The second clade comprised U. tigridis samples from Israel ( Lake Kinneret in the Jordan River basin) and Turkey (the Tersakan stream) ( Fig. 2 View Figure 2 ). The M-PTP analysis considered these samples as two different species ( Fig. 1 View Figure 1 ), with a genetic distance of 2.89% ( Table 1 View Table 1 ). Only six specimens were analysed from this area, and the two haplotypes detected were separated by 32 substitutions (Supporting Information, Fig. S5 View Figure 5 ).
This species is typified by a short shell with anteriorly shifted umbos, which was observed in the Israeli specimens. The Turkish specimens resembled the U. mancus morph ( Fig. 6 View Figure 6 ).
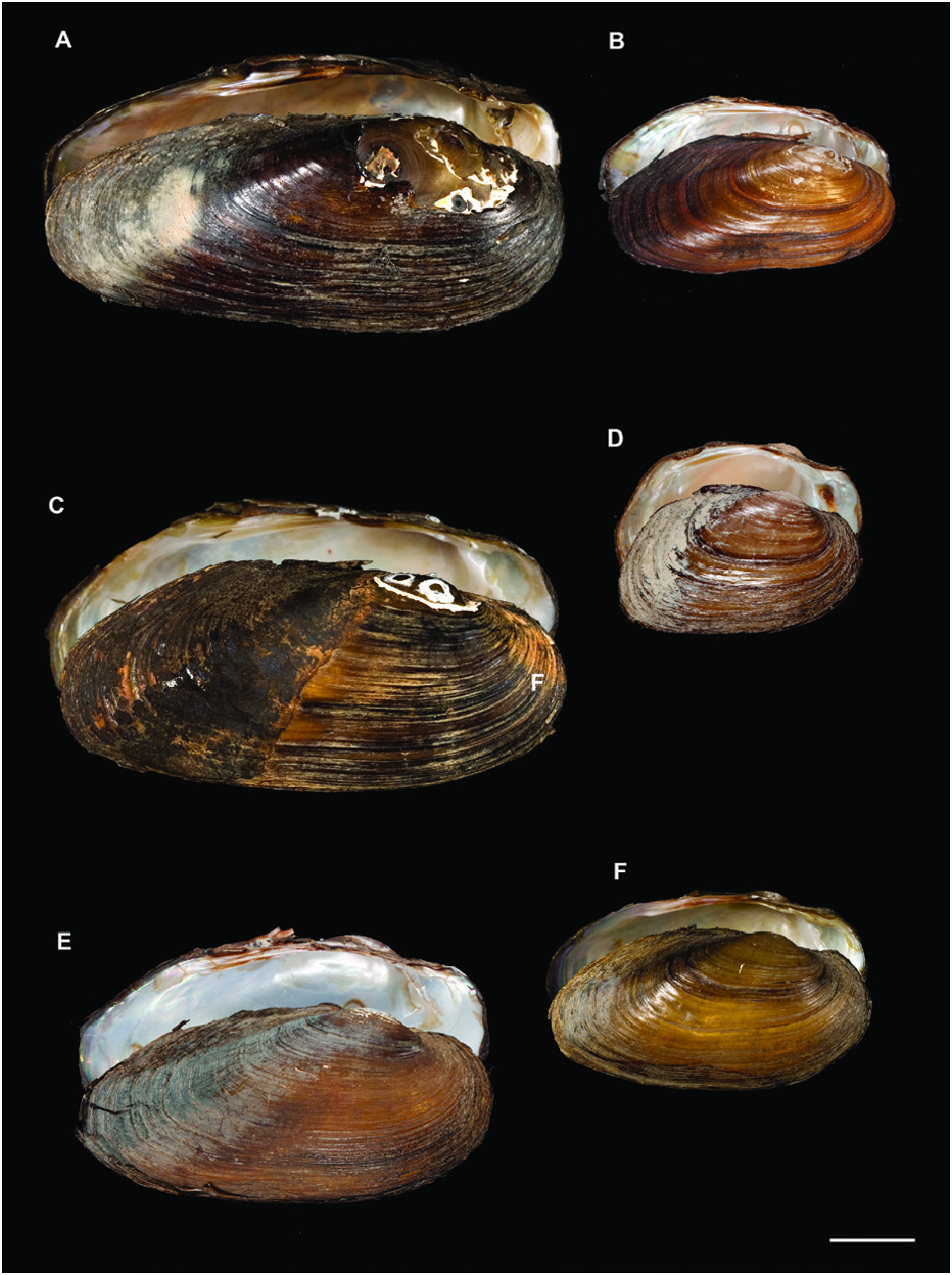
The next lineage combined the U. ravoisieri specimens into two subclades with a genetic distance of 2.55% ( Table 1 View Table 1 ; Fig. 2 View Figure 2 ). These subclades were also consistently recovered as distinct species in the bGMYC and M-PTP analyses. The two most frequent haplotypes (Supporting Information, Fig. S6 View Figure 6 ) were separated by 18 steps. One of these haplotypes represented the Spanish samples, and was very close to a haplotype present in the Tunisian Kebir River (separated by only one step), and to one of the two divergent El Maaden haplotypes (separated by four steps).
The shell of this species typically has a modified U. mancus shape, but the morphs we obtained were highly variable, especially those from the Spanish Ser River ( Khalloufi et al., 2011: fig. 3).
The clade consisting of U. mancus ( Fig. 1 View Figure 1 ) included specimens from Mediterranean basins in Spain (the
The intraspecific divergences within species divided into different units were Unio crassus : 2.10, Unio pictorum : 0.72, Unio ravoisieri : 1.36, Unio tigridis : 0.96 and Unio gibbus : 1.18%.
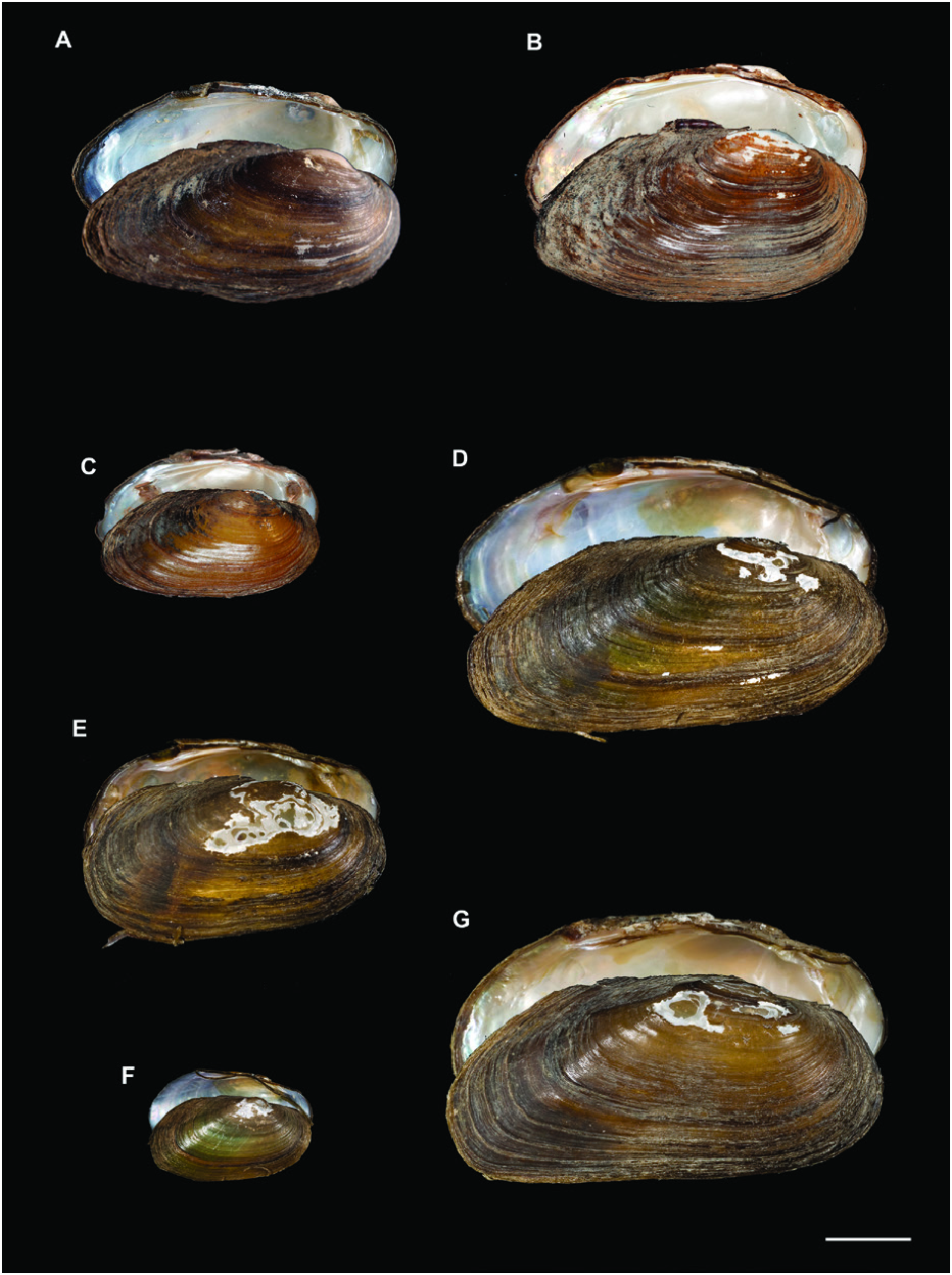
Ebro, Fluviá, Ter, Sonella, Llobregat and Júcar rivers), France (the Hérault and Argens rivers, the Tet River in the Basse Basin, Lake Bourget and the Ognon River, both within the Rhône Basin, many rivers in Corsica and the Atlantic rivers Charente, Seine and Drée, a tributary of the Loire) and Italy (many rivers in Sardinia) ( Fig. 2 View Figure 2 ). The network analysis showed haplotypes with some biogeographic incongruences (Supporting Information, Fig. S7 View Figure 7 ). For instance, the Júcar and Sonella rivers (eastern Spain) shared a haplotype with some French populations, but not with any of the northeastern Spanish populations. This high-frequency haplotype appeared in both Atlantic and Mediterranean French and Spanish rivers. Of the 15 different haplotypes found, five were exclusive to Sardinian-Corsican populations, one to the French Lake Bourget and one appeared with a high frequency in the Catalonian (eastern Spain) rivers.
This species had a more rounded shell than the other Mediterranean species U. elongatulus , with the exception of specimens from northeastern Spain, Corsica and Sardinia, which were elongated ( Figs 7 View Figure 7 , 8 View Figure 8 ).
The next clade included the U. elongatulus ( Fig. 1 View Figure 1 ) specimens from Mediterranean freshwaters in Italy (Po and Isonzo rivers and Alpine lakes), Croatia (Mirna and Zrmanja rivers, Bačinska Lake) and Albania (Scutari Lake) ( Fig. 2 View Figure 2 ). The Croatian haplotypes appeared at both ends of the network (Supporting Information, Fig. S8 View Figure 8 ), therefore, shared haplotypes between Croatia and Italy would be expected. However, no haplotypes were shared among any of the specimens from these regions.
The specimens had more elongated shells than other Mediterranean species ( U. mancus ) and a posterior umbo, although some populations had more rounded shells ( Fig. 9 View Figure 9 ).
The last clade represented the species U. pictorum , which included two groups having a genetic distance of 2.38% ( Table 1 View Table 1 ). The first group (1) consisted of specimens from Western and Central Europe (Lake Bourget in the Rhône Basin and the Adour, Seine, Oder, Danube and Thames basins), eastern Greece (Strymonas and Axios rivers and Lake Volvi), Ukraine (Teteriv at the Black Sea), Iran and Russia (Dzhankhot River at the Black Sea, Vyborg River in Karelia). The second group (2) comprised specimens from both western (Trichonida Lake at the Acheloos basin) and eastern (Axios River) Greece ( Fig. 2 View Figure 2 ). These two subclades were consistently recovered as distinct species in the bGMYC and M-PTP analyses. Up to 16 different haplotypes (Supporting Information, Fig. S9 View Figure 9 ), with from one to a maximum of 23 substitutions, were found among the specimens of this species. There was a maximum of 8 steps of differentiation within group ‘1’ and 17 between two of the three ‘2’ haplotypes. The network shape of group ‘1’ represented polymorphic populations with two relatively frequent haplotypes, one of which was only found in specimens from the Thames River and the other shared by samples as far away as Iran and Greece. Haplotypes found in the Danube and Dzhankhot rivers and in Lake Volvi seemed to derive from the latter one.
This highly polymorphic species had a typical elongated shell. However, some specimens were very different, for example those from Strymonas River, Lake Volvi and the Axios River (within both clades). Some populations resembled U. mancus (Lake Volvi, Thichonida and Strymonas rivers) ( Figs 10 View Figure 10 , 11 View Figure 11 ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.