
Bonita mexicana, Campos, Ernesto, 2009
|
publication ID |
https://doi.org/10.5281/zenodo.186086 |
|
DOI |
https://doi.org/10.5281/zenodo.6222594 |
|
persistent identifier |
https://treatment.plazi.org/id/743587B5-FFC1-FFF9-19BF-2686FC46F92D |
|
treatment provided by |
Plazi |
|
scientific name |
Bonita mexicana |
| status |
sp. nov. |
Bonita mexicana View in CoL new species
( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 B, 8B)
Distribution and host. Known only from the type locality, Point Sofia, Tortugas Bay, Baja California Sur, México ( 27º 41.943’ N; 114º 52.542’W). Found in the mantle cavity of Pseudochama exogyra (Conrad, 1837) (Mollusca, Pelecypoda, Family Chamidae ) cemented to rocky reef.
Type material. Female holotype, (CNCR-25300), 6 females paratypes (CNCR-25301, USNM), 26–27 Dec 1989.
Other material examined. 1 female ( UABC), Dec. 1990; 3 females ( UABC), 2 Jan 1992; 7 females ( UABC), 2–3 Dec. 1994, Point Sofia, Tortugas Bay, Baja California Sur, México.
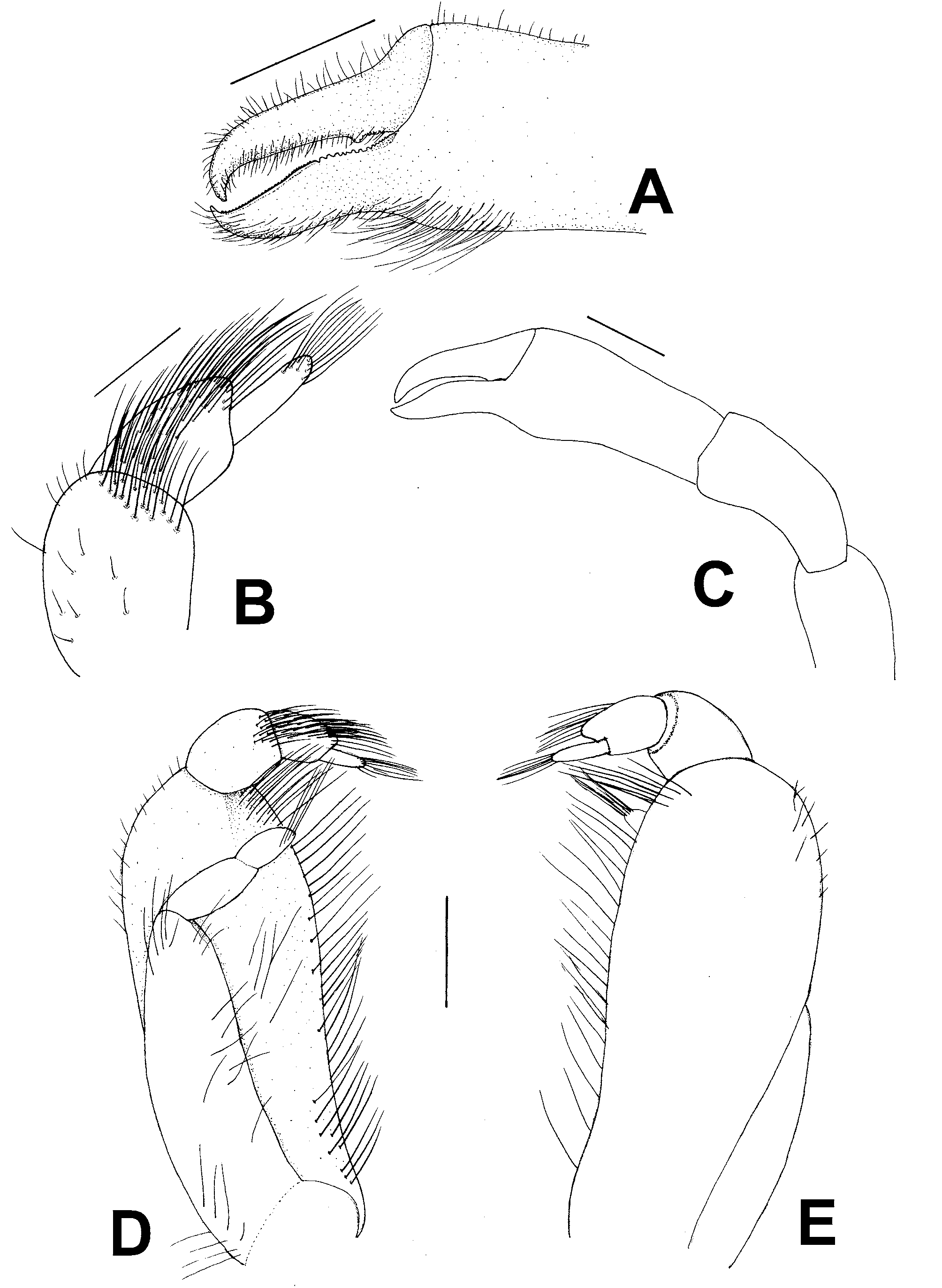
Description. Carapace subhexagonal, tumid, regions well defined, smooth, shiny, anterolateral margins protruding beyond frontal margin; 2 longitudinal, deep sulci extending form orbits to middle of carapace; posterior margin somewhat sinuous ( Fig. 2 View FIGURE 2 B). Orbits small, eyes oval, visible in dorsal view. Prominent protuberance at posterior end of basal segment of antenna ( Fig. 2 View FIGURE 2 A). MXP3 obliquely placed on buccal cavity, ischium, merus indistinguishably fused, inner, outer margin gently curved, lacking obtuse angle ( Fig. 3 View FIGURE 3 D–E); palp 3-segmented; carpus larger than propodus, latter extending distally, dactylus digitiform distally inserted on distoventral angle of propodus ( Fig. 3 View FIGURE 3 B). Exopod of MXP3 with 2-segmented flagellum ( Fig. 3 View FIGURE 3 D).
Chelipeds elongated ( Fig. 3 View FIGURE 3 C), chelae subcylindrical; manus increasing in width distally; pollex slightly deflexed, ventral margin proximally concave, distally convex; dactylus wide at base, distally with sharphooked, hard tip; cutting edge of dactylus with small proximal tooth, both fingers with additional minute teeth ( Fig. 3 View FIGURE 3 A).
Walking legs slender, relative length 3> 2> 1> 4; WL3 asymmetrical, left 1.2 times longer than right ( Fig. 2 View FIGURE 2 C). Propodi with margins subparallel except those of WL4 distally expanded. Dactyli curved at tip, relative length 3> 2> 4> 1, those of longer WL3 blade-shaped.
Abdomen greatly enlarged, with 6 somites and telson well separated ( Fig. 4 View FIGURE 4 A), strongly concave, laterally covering ischia, margin with tomentum, involuted, ( Fig. 4 View FIGURE 4 B–C), distally covering the buccal cavity.
Male. Unknown.
Etymology. The name mexicana refers to the country where the type material was collected.
Remarks. The soft carapace, the protuberance on the basal article of the antennae, the transversely placed MXP3 on the buccal cavity, the slender and feeble walking legs and the well-separated six-segmented abdomen and telson are features shared between Bonita and the genera Arcotheres Manning, 1993 (see Campos & Manning, 2001), Austinotheres Campos, 2002 , Epulotheres Manning, 1993 , Gemmotheres Campos, 1996 , Nannotheres Manning & Felder, 1996 , Orthotheres Sakai, 1969 , Ostracotheres H. Milne- Edwards, 1853, Pinnotheres Bosc, 1802 , Raytheres Campos, 2002 , and Zaops Rathbun, 1900 . However, the presence of a thin, translucent, easily wrinkled carapace and the enlarged MXP3 carpus suggest that Bonita is allied to the eastern Pacific genera Austinotheres and Enigmatheres . In contrast, the remarkably asymmetrical and enlarged WL3 is a character shared with the American genus Epulotheres Manning, 1993 and the Indo- West Pacific genus Arcotheres Manning, 1993 (see Manning, 1993b; Manning & Felder, 1996 and Campos & Manning, 2001). The remaining genera of the subfamily Pinnotherinae have symmetrical walking legs or asymmetrical WL2. Bonita can be easily distinguished from all the above genera by the presence of two sulci on the carapace, the anterolateral margins of carapace protrude beyond the frontal margin and by the clavate WL4 propodus. The two deep sulci on the carapace in Bonita are also present in members of the genus Fabia Dana, 1851 ( Fig. 5 View FIGURE 5 ). This feature may suggest that these two genera are allied. However, the genus Fabia differ from Bonita in the anterolateral margins of the carapace, that do not protrude beyond the frontal margin, the WL2 that are the longest, the propodi of the WL4 with their margins subparallel and the MXP3 with a propodus larger that the carpus and the dactylus inserting in an angular notch in the ventral middle third of the propodus (see Campos, 1996b; Fig. 2–3 View FIGURE 2 View FIGURE 3 vs. 6 A–B). These differences support the sulci of the carapace should be regarded as convergences. Noticeably is that the larval zoeal morphology of Bonita , particularly the shapes of the abdomen and telson, are similar to that of species of Arcotheres , Austinotheres , Gemmotheres , Nepinnotheres , Orthotheres , Ostracotheres , and Pinnotheres ( Fig. 8 View FIGURE 8 ). In contrast, the abdomen and telson in larval zoeal of Bonita markedly differ from those observed in the genera Fabia / Juxtafabia and Pinnixa ( Fig. 7 View FIGURE 7 ). These latter genera shared some larval features of the abdominal somites (see Marquez and Phole, 1995) which have recently been regarded as convergence of two independent monophyletic groups (see, Palacios- Thiel, et al., 2008). I consider Bonita and their allied genera represent another monophyletic group within the Pinnotheridae , as strongly suggest the shared adult and larval morphology herein discussed as well as the molecular evidence recently provided by Palacios-Thiel, et al., (2008).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |