
Myrmozercon karajensis, Joharchi, Omid, Halliday, Bruce, Saboori, Alireza & Kamali, Karim, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.202824 |
|
DOI |
https://doi.org/10.5281/zenodo.5668788 |
|
persistent identifier |
https://treatment.plazi.org/id/730FD955-FFED-FF82-4EF4-FC79FEA1FE04 |
|
treatment provided by |
Plazi |
|
scientific name |
Myrmozercon karajensis |
| status |
sp. nov. |
Myrmozercon karajensis sp. nov.
( Figures 19–27 View FIGURES 19 – 27 )
Specimens examined. Holotype, female, Iran, Karaj, 35˚55’ N, 51˚04’ E, alt. 1880 m, 15 April 2010, O. Joharchi coll., in nest of Camponotus sp. (in JAZM). Paratype, one female, same data as holotype (in ANIC).
Description. Female
Dorsal idiosoma ( Fig. 19 View FIGURES 19 – 27 ). Length 583–622 µm. Dorsal shield length 535–538 µm, width 491–504 µm (n = 2). Shield posteriorly truncate, not covering entire idiosoma, leaving a curved strip of unprotected skin posterior to setae J5, shield with distinct reticulate ornamentation over whole surface, individual polygons small, scale-shaped, transverse; with approximately 33 pairs of smooth pointed setae, some unpaired and asymmetrical, two pairs of setae in soft skin posterior to shield, setae on shield uniform in length and thickness except j1 fine and minute. Setae in R series reduced to only one or two setae. Pores and lyrifissures minute and obscure, in approximately ten pairs.
Ventral idiosoma ( Fig. 20 View FIGURES 19 – 27 ). Tritosternum with short broad base (37µm × 12 µm wide), with smooth paired laciniae (105 µm); pre-sternal shields absent. Sternal shield (length 124 µm) narrowest between coxae II (131 µm) widest between coxae II and III (178 µm), with biconvex anterior margin; posterior margin indistinct and concave; shield bearing three pairs of smooth pointed setae (st 1 40 µm, st 2 42 µm, st 3 40 µm) and two pairs of lyrifissures, one pair between setae st 1 and st 2 and the other between st 2 and st 3; surface with indistinct reticulate ornamentation and weak transverse lines near the anterior margin. Seta st 4 present in soft skin, metasternal pores and plates apparently absent. Endopodal plates III/IV free, narrow and elongate. Genital shield wide, strongly tapering posteriorly, 330–342 µm long, 150–153 maximum width. Surface of shield reticulate, ornamented with elongate cells in posterior half and longitudinal markings in anterior half; with one pair of simple setae st 5. Anal shield indistinct, bearing long thick post-anal seta 74 µm long, and a pair of para-anal setae 45 µm long. Opisthogastric soft skin with 11 pairs of smooth pointed setae (Jv1 32 µm, Jv2 37 µm, Jv3 17 µm, Jv5 44 µm, Zv1 37 µm, Zv2 37 µm), metapodal shields absent. Peritremes very short (71 µm), extending to mid-level of coxae Ш. Peritrematal shields absent, post-stigmatal plates and pores absent.
Gnathosoma . Hypostomal groove with ten rows of denticles, 9 to 15 very fine denticles per row, three rows sloping, not transverse (not visible in paratype) ( Fig. 21 View FIGURES 19 – 27 ). Hypostomal seta h1 26 µm, h2 23 µm, h3 37 µm, palp coxal seta 34 µm, surface of hypostome ornamented with transverse and curved lines. Palp chaetotaxy: trochanter 2, femur 5, genu 5, tibia 12; pd on femur very long and thick; palp tarsal claw two-tined. Epistome triangular, smooth, with pointed apex and a few longitudinal dorsal lines ( Fig. 22 View FIGURES 19 – 27 ). Fixed digit of chelicera edentate, longer than movable digit; movable digit weakly sclerotised, distally curved, with a row of about eight minute denticles ( Fig. 23 View FIGURES 19 – 27 ). Corniculi short, broad, weakly sclerotised.
Legs: Legs II and III short (465 µm, 505 µm), I and IV longer (623–703 µm, 314–347 µm), ventral surface of most segments with reticulate ornamentation. Chaetotaxy: Leg I: coxa 0 0/1 0/1 0, trochanter 1 0/1 1/2 1 (pd very long and thick), femur 2 3/1 2/1 2 (ad 1 and pd 1 long and thick, Fig. 24 View FIGURES 19 – 27 ), genu 2 3/1 2/1 2, tibia 2 3/1 2/1 2. Leg II: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 1 2/1 2/2 1 (ad 1 and pd 1 long and thick, Fig. 25 View FIGURES 19 – 27 ), genu 2 3/1 2/1 2, tibia 2 2/1 2/1 2. Leg III: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 2/1 1/0 1 (pd and pl very short and fine, ad 1 and ad2 long and thick, av proximal to lyrifissure, Fig. 26 View FIGURES 19 – 27 ), genu 2 2/1 2/1 2, tibia 2 1/1 2/1 2. Leg IV: coxa 0 0/1 0/0 0 (av very long and fine), trochanter 1 1/2 1/1 0, femur 2 2/1 1/0 1 (pd and pl very short and fine, ad 1 long and thick, Fig. 27 View FIGURES 19 – 27 ), genu 2 2/1 3/0 2, tibia 2 1/1 3/1 2. Tarsi I–IV with 16 setae, pre-tarsi with membranous ambulacrum, claws absent.
Insemination structures: Insemination ducts opening on posterior margin of coxa III; sacculus foemineus not visible, apparently unsclerotised.
Etymology. The name of this species is derived from the type locality, Karaj.
Notes. Only eight species of Myrmozercon have been described from the Palaearctic Region. Myrmozercon karajensis is distinctive in having only two ventral setae on genu I and tibia I instead of the normal three. It differs from M. brevipes Berlese, 1902 ( Italy) and M. flexuosa ( Michael, 1891) ( Austria) because it has many fewer setae on the dorsal shield; it differs from M. acuminatus ( Berlese, 1903, Italy) by its very short peritreme—in M. acuminatus the peritreme is long enough to reach coxa I ( Berlese, 1903). Myrmozercon karajensis differs from M. antennophoroides ( Berlese, 1904) ( Italy) because its genital shield is posteriorly pointed whereas that of M. antennophoroides is truncate; it differs from M. brachiatus ( Berlese, 1903) ( Italy) by the posterior margin of the dorsal shield, which is truncate in M. karajensis and rounded in M. brachiatus . Myrmozercon ovatum Karawajew, 1909 was described from Turkmenistan, close to the type locality of M. karajensis . However, M. ovatum is easily distinguished from M. karajensis by its very distinctive short legs, shorter than the width of the idiosoma, and its wide genital shield. Myrmozercon karajensis is most similar to M. diplogenius ( Berlese, 1903) ( Italy and East Africa), but differs from it because the posterior half of the dorsal shield has at least 19 pairs of setae in M diplogenius and 15 pairs in M. karajensis , and the posterior dorsal shield setae are 48–50 µm long in M diplogenius and 53–58 µm long in M. karajensis . Myrmozercon yemeni ( Ueckermann & Loots, 1995) was described from Yemen, geographically close to Iran, but it differs from M. karajensis in many ways, especially in the absence of metasternal seta st4, and the fusion of the sternal and endopodal shields. Myrmozercon tauricus Trach & Khaustov, 2011 , described from Ukraine, also lacks metasternal setae, and differs from M. karajensis in having pilose setae on the dorsal shield. Myrmozercon karajensis is very similar to M. chapmani ( Baker & Strandtmann, 1948) ( Mexico) , but differs from it by the presence of very long thick setae on the femora of all legs (absent in M. chapmani ).
Shaw & Seeman (2009) drew attention to some distinctive character states in Myrmozercon . The palp trochanter usually has one ventral seta, but M. karajensis and M. beardae have the normal mesostigmatid complement of two ventral setae. The corniculi of most species, including M. karajensis , are short and broad, not strongly sclerotised and horn-like. The genus is variable in the presence or absence of seta st 4 (present in M. karajensis ) and in the degree of fusion of the sternal shield with the endopodal plates. In M. karajensis the endopodal plates between coxae III and IV are completely free. Myrmozercon also has reduced chaetotaxy on some leg segments. This trend is developed further in Myrmozercon karajensis , which has only two ventral setae on genu I and tibia I instead of the normal three. This requires an expansion of the definition of the genus, and of the subfamily Melittiphinae.
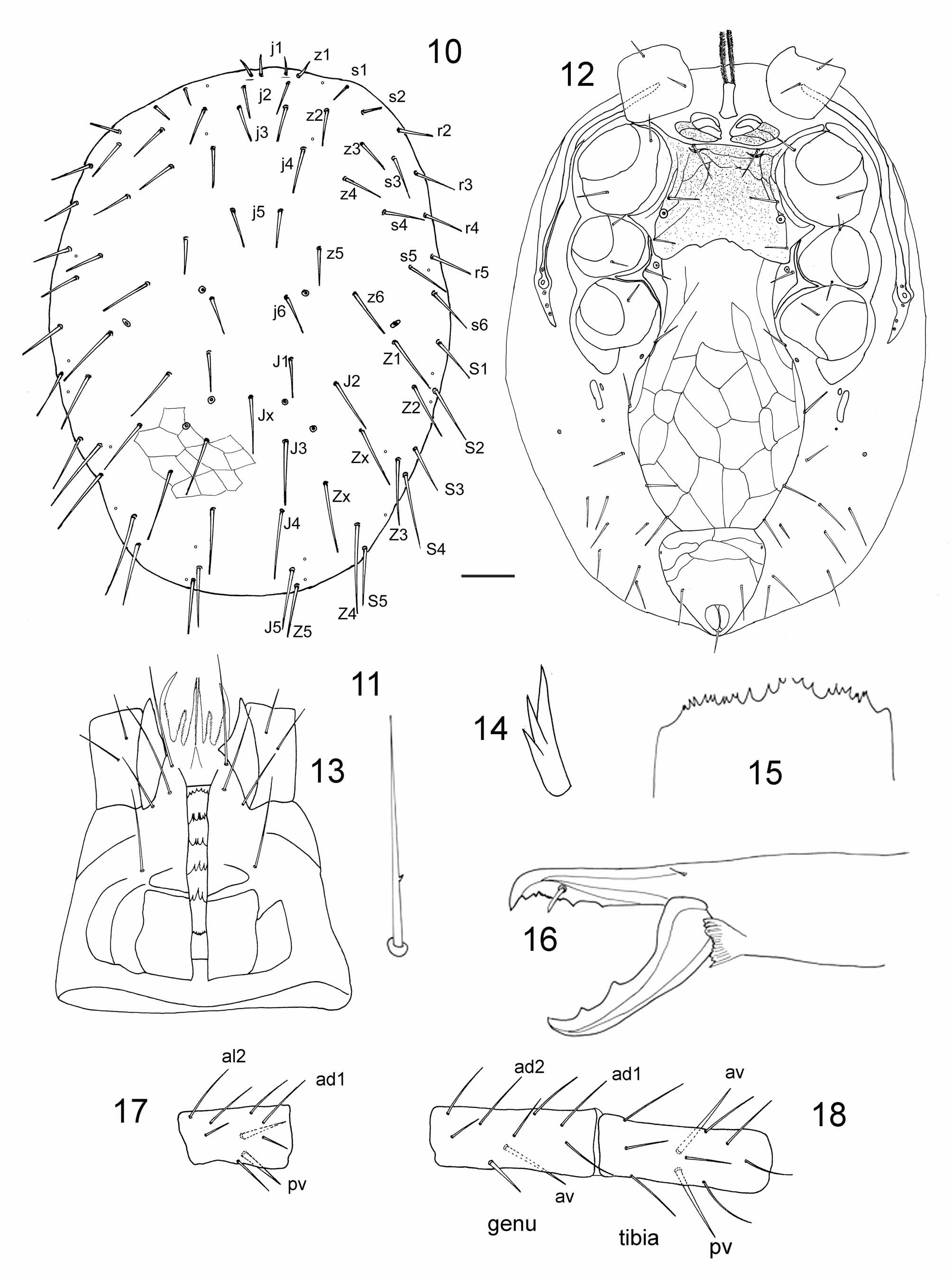
Most species of Myrmozercon , including the type species M. brevipes , show moderate to strong hypertrichy on the dorsal shield. However, M. burwelli Shaw & Seeman, 2009 (24–25 pairs), and the new species M. karajensis (33 pairs), have a reduced dorsal chaetotaxy. All species appear to have asymmetrical and unpaired setae on the dorsal shield, which makes it difficult to recognise their homology. For that reason, the setal notations in Figure 13 View FIGURES 10 – 18 must be regarded as provisional. In most species the dorsal shield is reduced or truncated posteriorly to expose a strip of unsclerotised opisthonotal skin, but this is not true in every species. Species of Myrmozercon also vary in the presence or absence of metasternal setae st 4, and the extent to which the sternal shield is fused with the endopodal plates. The sternal shield of M. karajensis is similar to that of many free-living Mesostigmata , with three pairs of setae and two pairs of lyrifissures, separate endopodal plates between coxae III and IV, and separate metasternal setae in the soft skin. The leg chaetotaxy of Myrmozercon species is variable, and does not provide diagnostic characters that define the genus ( Shaw & Seeman, 2009). The new species M. karajensis adds some further variation to the leg chaetotaxy, by having only two ventral setae on genu I and tibia I, where most species have three. It also has two ventral setae on the palp trochanter where most species of Myrmozercon have one. Shaw & Seeman (2009) described a swelling on the dorso-distal edge of the palp trochanter in several species, but this structure is not present in M. karajensis . This instability in morphology, and the edentate chelicerae and short peritremes of Myrmozercon , suggest that Myrmozeron is parasitic on its ant hosts, and not simply a commensal in its host's nests, but this has not been established experimentally. The specimens of M. karajensis were found clinging to the abdomen and head of the ants.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.