
Tsingymantis antitra, Glaw, Frank, Hoegg, Simone & Vences, Miguel, 2006
|
publication ID |
https://doi.org/ 10.5281/zenodo.174270 |
|
DOI |
https://doi.org/10.5281/zenodo.5668690 |
|
persistent identifier |
https://treatment.plazi.org/id/72204745-6757-FFFA-F64A-FAD0FD6FCBB6 |
|
treatment provided by |
Plazi |
|
scientific name |
Tsingymantis antitra |
| status |
sp. nov. |
Tsingymantis antitra View in CoL sp. nov.
Holotype. ZSM 304/2004 (fieldnumber FGZC 589), collected on 27 February 2004 below the "Point de Vue Petit Tsingy", 12°57'25''S, 49°07'06''E, 117 m alt, Ankarana Special Reserve, northern Madagascar, by F. Glaw, M. Puente & R. D. Randrianiaina.
Paratypes. ZFMK 84436 (originally ZSM 305/2004), same data as holotype; ZSM 769/2003 (fieldnumber FG/MV 2002-0577), cleared and stained specimen, collected on 12 February 2003, below "Campement des Anglais" (now called Campement Anilotra), Ankarana Special Reserve, northern Madagascar, by F. Glaw, R. D. Randrianiaina & A. Razafimanantsoa; UADBA 24766 (fieldnumber FGZC 531), collected on 25 February 2004 close to the "Grotte des Chauve-souris", 12°57'S, 49°07'E, ca. 50 m alt, Ankarana Special Reserve, northern Madagascar, by F. Glaw, M. Puente & R. D. Randrianiaina.
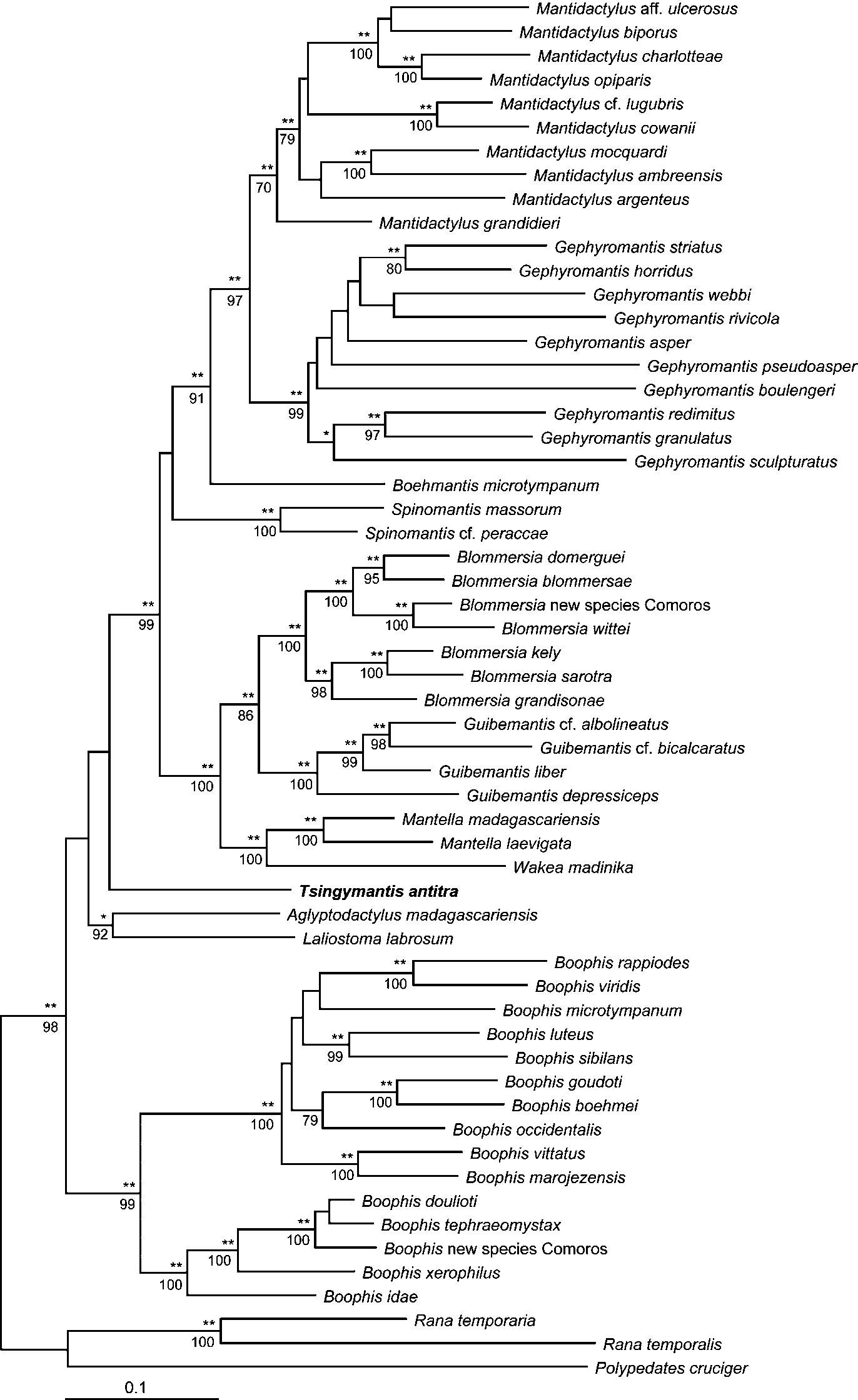
Diagnosis. A large species of mantellid frogs representing an isolated and basal lineage within the family, based on a molecular analysis of mitochondrial and nuclear genes ( Fig. 3 View FIGURE 3 ). Differs from all other mantellid genera and subgenera as outlined in the diagnosis of the genus Tsingymantis . It differs from all other large mantelline and laliostomine species that reach a SVL of more than 60 mm as follows: From Mantidactylus mocquardi , M. grandidieri , M. guttulatus , M. ambohimitombi , Boehmantis microtympanum , and Aglyptodactylus madagascariensis by largely connected lateral metatarsalia (versus separated), from Aglyptodactylus laticeps and Laliostoma labrosum by distinctly enlarged tips of fingers and toes, by the presence of a complete circummarginal groove on pads of fingers and toes, and by having the first finger shorter than the second.
Description of the holotype. Adult female (with oocytes in the body cavity), in good state of preservation but with a midventral slit. SVL 67.1 mm, for further measurements see table 1. Body slender; head wider than body; snout approximately rounded in dorsal and lateral views, nostrils directed laterally, protuberant, much nearer to tip of snout than to eye; canthus rostralis distinct, straight; loreal region concave; tympanum very distinct, rounded, 76 % of eye diameter; supratympanic fold distinct, curved; tongue was ovoid and bifid posteriorly (part of the tongue was removed as DNA sample); vomerine teeth present in two groups, maxillary teeth present; choanae relatively rounded. Arms slender, subarticular tubercles single; fingers without webbing; relative length of fingers 1<2<4<3; finger disks distinctly enlarged; nuptial pads absent. Hind limbs slender; tibiotarsal articulation reaches the eye when the hind limb is adpressed along the body; lateral metatarsalia largely connected; inner metatarsal tubercle distinct, outer metatarsal tubercle absent; webbing formula (according to Blommers-Schlösser 1979) between toes 1(1), 2i (1), 2e(0.5), 3i (1.5), 3e(1), 4i (2.5), 4e(2), 5(1); relative length of toes 1<2<3<5<4. Skin on the upper surface smooth, without folds or ridges. No distinct enlarged tubercles in the cloacal region; ventral skin smooth, finely granular on the shanks. No femoral glands.
Colouration of the holotype. After 1.5 years in preservative, back blackish with indistinct dark brown reticulations and a few small greyish spots above the insertion of the left arm. Upper surfaces of arms and legs dark brown with indistinct black markings. Flanks lighter brown than back, although there is no distinct colour border between flanks and back. Tympanum light brown in periphery, darker brown in center. Ventrally dirty cream-whitish on belly, with fine indistinct mottling. Throat brown, chest and ventral surfaces of arms light brown, ventral surface of hindlegs yellowish in the center becoming darker brown to periphery. Ventral side of lower leg and tarsus dark brown, foot and webbing brown.
The dorsal ground colour in life was brown with violet shade and with olive green spots ( Fig. 1 View FIGURE 1 ). The iris was silvery-grey with a brownish horizontal streak and a bluish iris periphery. The ventral surface was pinkish to brown ( Fig. 2 View FIGURE 2 ).
Variation (see table 1 for measurements). The ZFMK and ZSM paratypes are very similar to the holotype in size and morphology. The colouration of ZFMK 84436 is generally similar to that of the holotype, but sligthly lighter, that of ZSM 769/2003 (assessed before clearing and staining) distinctly lighter, especially on the ventral side. All three specimens have pigmented oocytes in the body cavity although they are relatively small in size and numbers. The UADBA paratype is similar to the other specimens in size and colour, but was not available for detailed studies.
Osteological features. A number of skeletal characters known to be relevant in mantellid systematics were assessed on the cleared and stained paratype ZSM 769/2003. Maxillary and vomerine teeth present. Omosternum forked, the greatest space between the arms being about two times the width of one arm, sternum unforked, bony part of sternum longer than that of omosternum. Hyoid with a distinct anterolateral and a small posterolateral process. Intercalary element present between ultimate and penultimate phalanges of all fingers and toes. Terminal phalanges distinctly Y-shaped, with rather broad and posteriorly serrated arms. Three free distal tarsals, the third tarsal being small.
Etymology. The specific name is derived from the Malagasy word " antitra " (meaning old) and refers to the presumed old age of the Tsingymantis lineage. The name is considered as an invariable noun standing in apposition to the genus name.
Habitat and natural history. The Ankarana reserve consists mainly of bizarre, eroded tsingy limestone formations and moderately dry forest areas. It is crossed by four rivers and includes more than 100 km inventorized subterraneous passages and caves. The climate of Ankarana is "dry tropical" with a long dry season between May and December. The wettest months are January, February, and March. There are about 86–92 rainy days per year. The hottest month is March (36.2°C) and the coldest month is June (13.5°C) (http://www.parcs-madagascar.com/ankarana/index.htm, as of 20 August 2005). All four specimens of Tsingymantis antitra were exclusively found at night and associated with tsingy formations. ZSM 769/2003 was found along a small brook, whereas the other three specimens were not found associated with open waters (although not far from dry riverbeds). UADBA 24766 was sitting on the ground close to the entrance of a big cave. The two other specimens were found on the top of the tsingy limestone and in a cave of ca. 1 m depth in tsingy, respectively, both close to a dry riverbed. A further individual was photographed along a brook (Paul Freed, pers. comm.). However, it remains unclear if the species is mainly distributed along streams or widespread in the tsingy formations of Ankarana.
No unidentified frog calls were heard during the surveys and only small oocytes were found in the collected females, indicating that they were reproductively quiescent when collected. The period and mode of reproduction remain entirely unknown and at current no indications of cave breeding are known. ZSM 769/2003 had remains of a large orthopteran in the stomach.
Distribution and conservation status. Tsingymantis antitra is only known from the Ankarana Special Reserve in northern Madagascar which has a total surface of 182 km 2 ( Hawkins et al. 1990). The types of Tsingymantis antitra are from two areas in the Ankarana reserve, around the "Petit Tsingy" and around the "Campment des Anglais". The specimen photographed by P. Freed was discovered in a third area close to the "Campement des Americains" (now called "Campement d'Andrafiabe") in the west of the reserve, indicating that the species has a wider distribution in the central part of Ankarana. Connecting these three localities to a triangle allows to estimate the known extent of occurrence (see http://www.redlist.org/info/categories_criteria2001 for definition) for this species as much smaller than 100 km 2 (this approach was also used to calculate the extent of occurrence of the other Malagasy amphibian species by the Global Amphibian Assessment, see Andreone et al. 2005). The actual range is certainly larger, but since the tsingy-dependent fauna and flora of Ankarana includes many presumed local endemics (e.g., the snake Alluaudina mocquardi ) it appears likely that T. antitra is endemic to this reserve as well although it cannot be excluded that the species also occurs in other remote tsingy formations (e. g. Tsingy de Namoroka or Tsingy de Bemaraha) or other karstic areas (e. g. Analamera reserve). Due to its small assumed extent of occurrence (<182 km 2), its very small known extent of occurrence (<100 km 2), its very small known area of occupancy (<10 km 2), its apparent rareness (only four specimens have been found), and the fact that it represents a very ancient relict lineage we consider Tsingymantis antitra as "endangered" although its habitat currently seems to be relatively well protected.
Molecular phylogenetic relationships. The tree shown in Fig. 3 View FIGURE 3 includes for the first time representatives of all known lineages (genera, subgenera, and species groups) of mantellid frogs. Bayesian analysis of the molecular data yielded a tree ( Fig. 3 View FIGURE 3 ) with high support for most nodes. Especially the deeper nodes received much higher support than in the tree shown in Glaw & Vences (2006) which was obtained from less a complete data matrix. Maximum Bayesian support (100%) was found for all genera as defined in Glaw & Vences (2006), and for many subgenera and species groups. The included species of the Mantellinae (to the exclusion of Tsingymantis ) were highly supported as monophyletic group as well, with 100% Bayesian and 99% bootstrap support, and the same values were obtained for the species of the Boophinae . However, the relationships between the major mantellid lineages were not significantly resolved. The analysis placed Tsingymantis sister to the Mantellinae , and the Laliostominae ( Aglyptodactylus and Laliostoma ) sister to the Tsingymantis / Mantellinae clade. The Boophinae (genus Boophis ) occupied the most basal position. However, none of these groupings received Bayesian support of 95% or higher, or bootstrap support of 70% or higher, and the molecular data can therefore merely seen as weak indication of possible relationships among the major mantellid lineages. In any case, Tsingymantis has a very isolated position.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |