
Leptocheilopora, LANG, 1916
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00179.x |
|
persistent identifier |
https://treatment.plazi.org/id/6F0C243F-FF9F-254E-FC26-63B505D4F92E |
|
treatment provided by |
Diego |
|
scientific name |
Leptocheilopora |
| status |
|
GENUS LEPTOCHEILOPORA LANG, 1916 View in CoL LEPTOCHEILOPORA TENUILABROSA LANG, 1916
( FIGS 18A- C View Figure 18 , 22D View Figure 22 , 24O View Figure 24 )
Material: NHM D28892, Cretaceous , Santonian , Marsupites Zone , Upper Chalk , Brighton, Sussex, England . NHM D21210, Cretaceous , Santonian , Marsupites Zone , Upper Chalk , Roke Farm, south-east of Odiham, Hampshire, England . NHM D29901, Cretaceous , Santonian coranguinum Zone , Upper Chalk , Houndsdean Bottom , west of Lewes, Sussex, England .
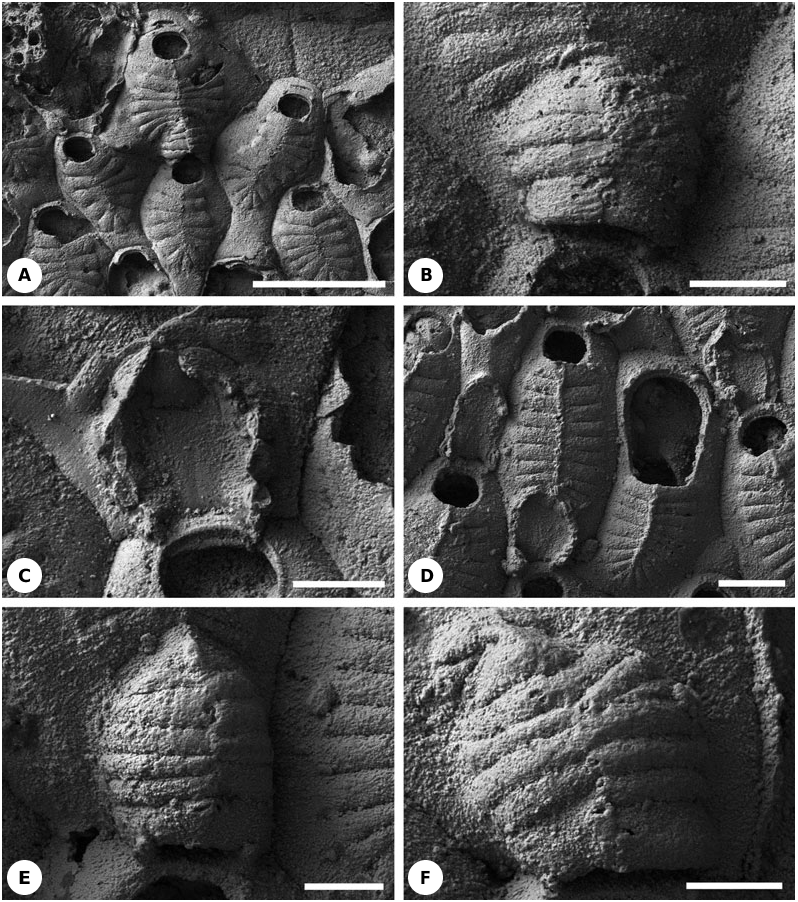
Description: Non-maternal zooids possess 2–4 small oral spines (four according to Lang, 1921 and Larwood, 1962). Occasionally two oral spines can be found in maternal autozooids. The frontal shield consists of 14–23 costae (14–15 according to Lang, 1921 and 17–22 according to Larwood, 1962) ( Fig. 18A View Figure 18 ) with brooding zooids having fewer costae than nonbrooding zooids. There is no evidence for the existence of the ‘intercostal spaces’ described by Larwood (1962: 107–108).
The hyperstomial ovicells are formed on the proximal gymnocyst of the distal (brooding) autozooid. Their external surface is distinctly costate, and they have a longitudinal medial suture that is aligned with the medial suture of the frontal shield of the distal zooid. Each ovicell roof consists of 11–14 tightly juxtaposed, centripetally orientated costae ( Figs 18A, B View Figure 18 , 23D View Figure 23 ). The bases of the costae are arranged in a semicircular (horseshoe) pattern ( Figs 18C View Figure 18 , 24O View Figure 24 ). Their distal tips meet and sometimes overlap on the midline of the ovicell roof. Ovicell costae have the same width as frontal shield costae, within the normal limits of variation. Distalmost ovicell costae are the shortest, often having a subtriangular shape, and are juxtaposed with the proximal (and shortest) frontal shield costae of the distal zooid. The two most proximal ovicell costae are located close to the distal boundary wall of the maternal zooid. The ovicell floor, surrounded by the costal bases, is slightly depressed. Larwood (1962: 109) described ‘occasional minute pores along... mid-lines’ of the ovicell costae but these have not been confirmed.
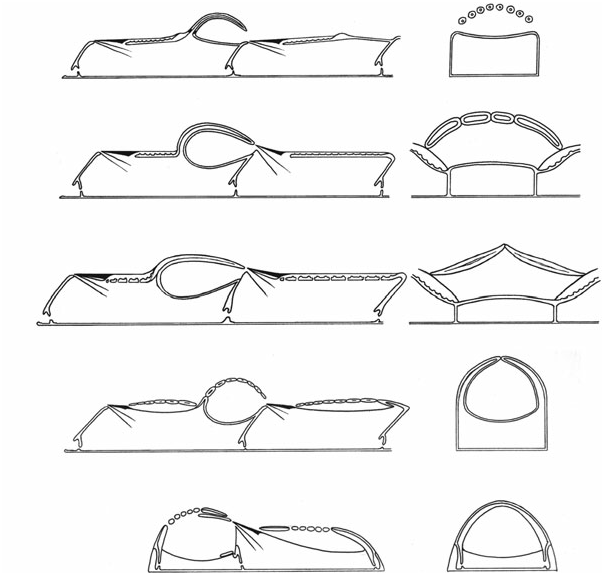
Costae of the only undamaged ovicell seen in specimen NHM D21210 expand and contract in width at intervals along their lengths in concert with neighbouring costae. The resultant wave-like form of the edges of the costae gives the impression that the costae fuse at intervals ( Fig. 18A View Figure 18 ; see also text-fig. 1B of Ostrovsky & Taylor, 2004).
Remarks: The first illustrated description of costate hyperstomial ovicells in any cheilostome was made for this species by Lang (1921). The same author had earlier (1916: 396) diagnosed L. tenuilabrosa in a species key as having ‘Ovicells formed like the intraterminal front wall’, indicating that he had already recognized their costal structure. When describing ovicells in his new species Cribrilina tumuliformis , which was subsequently placed by Lang (1921) in synonymy with L. tenuilabrosa, Brydone (1917: 51) remarked that ‘in a favourable light it can be discerned that these ooecia are ribbed like the zooecia.’ Larwood (1962) described and illustrated ovicells in L. tenuilabrosa , writing that ‘costae... cover the ovicell’. He was uncertain if ‘the costae may form an ectooecium’ (p. 109). Ryland (1979: 214) followed Larwood in suggesting that ‘the ooecium was either formed by or covered with a series of spines’, whereas Ostrovsky (2002) showed the ooecial wall to be formed entirely of costae.
( FIGS 18D- F View Figure 18 , 22D View Figure 22 , 24O View Figure 24 )
Material: NHM D55505, Cretaceous, Campanian, mucronata Zone, low in the Weybourne Chalk, Keswick Chalk Pit, near Norwich, Norfolk, England, P. Whittlesea Collection .
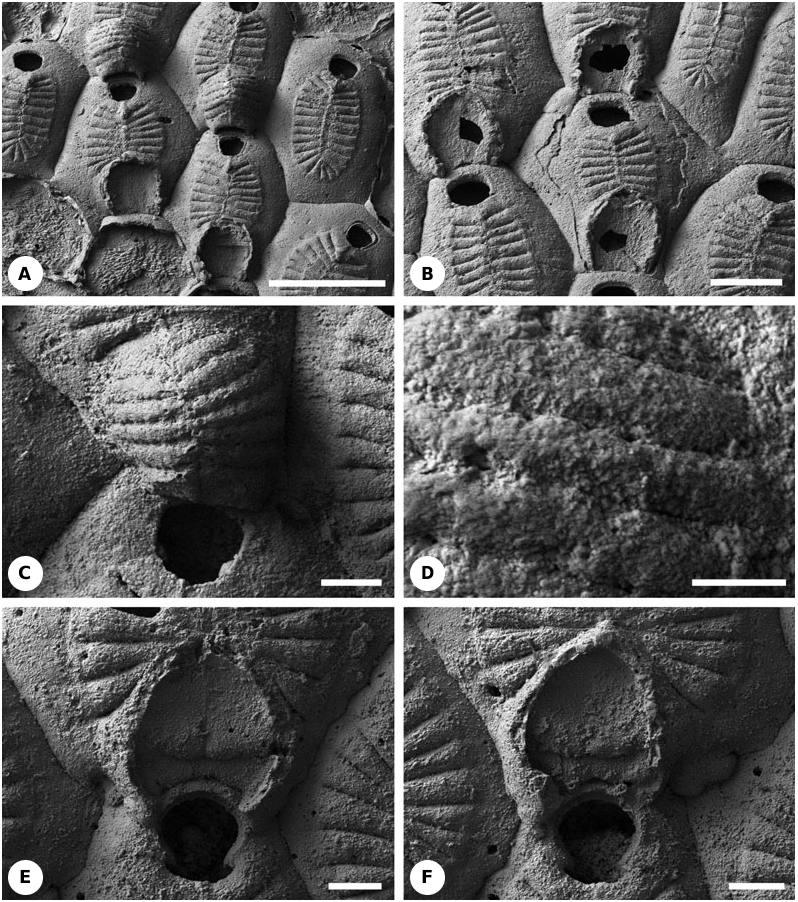
Description: All autozooids possess two small oral spines. In nonbrooding autozooids, the frontal shield comprises 21–22 costae whereas in brooding autozooids there are 15–19 costae, closely juxtaposed and without intervening slits ( Fig. 18D View Figure 18 ).
The hyperstomial ovicells are formed on the proximal gymnocyst of the distal brooding autozooid. Their roof is composed of 12–13 hollow juxtaposed costae, whose distal tips meet to form a low median keel which is aligned with the medial keel of the frontal shield ( Figs 18E, F View Figure 18 , 22D View Figure 22 ). Ovicell costal bases are arranged in a semicircular (horseshoe) pattern ( Figs 18D- F View Figure 18 , 24O View Figure 24 ). The two most proximal ovicell costae are located close to the distal boundary wall of the maternal zooid. The gymnocystal ovicell floor is almost flat or slightly concave ( Figs 18D View Figure 18 , 22D View Figure 22 ). Judging from the positions of occasional closure plates, which tend to be secreted immediately underneath the operculum in cheilostomes, the ovicell opening may not have been closed by the operculum during life (acleithral). Ovicell costae have approximately the same width as the frontal shelf costae. The two most proximal ovicell costae are the widest, whereas the most distal ovicell costae are the shortest. The latter juxtapose the proximalmost (also the shortest) costae of the frontal shield of the distal zooid. In one instance an ovicell is separated from the costate frontal shield of the brooding zooid by a substantial expanse of proximal gymnocyst ( Fig. 18D View Figure 18 ).
( FIGS 19A- D View Figure 19 , 22D View Figure 22 , 24O View Figure 24 )
Material: NHM BZ5204–6 , Cretaceous, Lower Maastrichtian , Porosphaera Beds, Sidestrand , Norfolk, England, P. Whittlesea Collection .
Description: There are no oral spines. Non-brooding zooids possess 21–29 costae and brooding zooids 15–19. The costae are tightly juxtaposed with their neighbours and have one or two small lumen pores (pelmatidia) near their distal ends ( Fig. 19A, B View Figure 19 ).
The hyperstomial ovicells are constructed of 14–15 juxtaposed costae on the proximal gymnocyst of the distal brooding zooid ( Figs 19A- D View Figure 19 , 22D View Figure 22 , 24O View Figure 24 ). One example has been found of an ovicell formed by an intramural reparative autozooid ( Fig. 19B View Figure 19 ). Distal ends of the overarching costae meet to form a median keel or sometimes a suture ( Fig. 19A, C View Figure 19 ). These structures are in alignment with the median keel of the frontal shield. Costal bases of the ovicells are arranged in a semicircular (horseshoe) pattern. The ovicell floor is slightly concave and is longitudinally elongated or equidimensional ( Fig. 19B View Figure 19 ). The two most proximal ovicell costae are positioned close to the distal boundary wall of the maternal zooid and are wider than the other costae. The distalmost ovicell costae are the shortest, often having a subtriangular shape. They are juxtaposed with the short, proximal costae of the frontal shield of the distal zooid. Ovicell costae are of roughly the same width as frontal shield costae.
As in the specimen (NHM D21210) of Leptocheilopora tenuilabrosa described above, junctions between ovicell costae have a wavy appearance, with intercostal sutures alternating with small pits ( Fig. 19C, D View Figure 19 ). This gives the appearance of lateral costal fusions, although there is a clear demarcation line (suture) between the seemingly fused parts of the costae.
Remarks: Larwood (1962) suggested that Leptocheilopora magna Lang, 1916 , with which this species can be compared, possessed costate ovicells. Unfortunately, there were no intact ovicells of L. magna at our disposal, although examination of broken ovicells in NHM BZ5207 suggests that Larwood’s opinion may be correct. The base of broken ovicells shows oval flattened elements, possibly broken costal bases, with an inverted T-shaped arrangement of closed slits ( Fig. 19E, F View Figure 19 ). This resembles the bilaterally symmetrical fold on the ovicell floor described in some Recent species of the cribrimorph Puellina ( Ostrovsky, 2002) .
Lang (1921) and Larwood (1962) recorded costate ovicells in the Cretaceous cribrimorphs Andriopora homunculus Lang, 1916 , Pliophloea striata Lang, 1916 , Aeolopora distincta Lang, 1916 and Castanopora dibleyi (Brydone, 1906) . We have reinvestigated the specimens described by these two authors using SEM and have been unable to find any trace of costae in the ovicells of any of these species.
Additional data from the literature: Voigt (1994) described and illustrated a new cribrimorph, Craticulacella schneemilchae Voigt, 1994 , from the Lower Campanian of Lägerdorf, near Hamburg, Germany. A broken costate ovicell is seen in Voigt’s plate 5, figure 1. It is represented by an estimated 8–10 costal bases arranged in a semicircular (horseshoe) pattern on the proximal gymnocyst of the distal zooid. The two most proximal ovicell costae are positioned close to the two oral spines, which are of similar size, and to the distal boundary wall of the maternal zooid. In contrast with species of Leptocheilopora , ovicell costal bases are spatially separated. Coupled with the fact that the costae forming the frontal shield have open slits between them, this feature suggests that intact ovicells probably had intercostal gaps, at least around the edges of the ovicell.
A similar ovicell structure is found in the undescribed cribrimorph? Thoracopora sp. (Lower Cenomanian, Mülheim, Germany), illustrated by Ostrovsky & Taylor (2004: text-fig. 1C). A half-broken ovicell has seven flat costae borne on the proximal gymnocyst of the distal zooid. Preserved parts of the costae are juxtaposed, but there are distinct sutures between them that may possibly be very narrow slits. An interesting point is that there are no distal (medial) spine bases in the two other, almost completely broken ovicells in this specimen. Thus, the ovicells of? Thoracopora sp. were probably formed by two lateral rows of costal bases. This may also be true for Craticulacella schneemilchae ( Fig. 24N View Figure 24 ).
Judging from the illustrations, costate ovicells may also occur in the cribrimorph Lekythoglena ampullacea Marsson, 1887 (pl. 9, fig. 7) but this needs to be confirmed using SEM.
GENUS BELLULOPORA LAGAAIJ, 1963 View in CoL BELLULOPORA BELLULA ( OSBURN, 1950) View in CoL
( FIGS 20A- F View Figure 20 , 22E View Figure 22 )
Material: NHM 1986.8.14.20, Recent, Florida, Atlantic Ocean, Station GOS 736, depth 60–90 m.
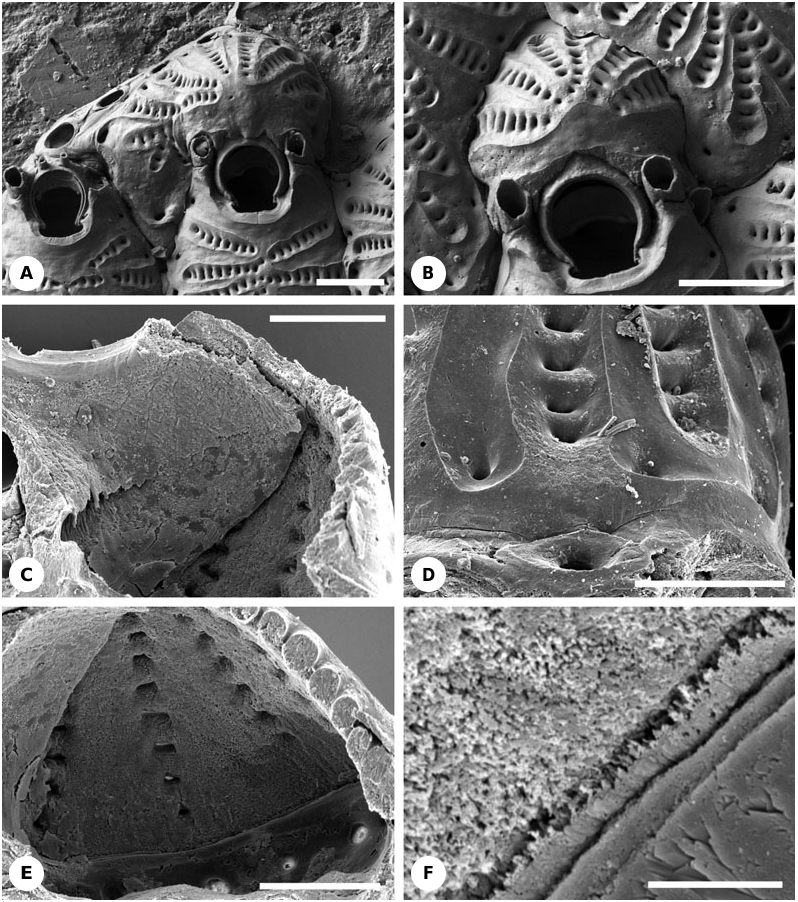
Description: Non-ovicellate autozooids have only one or two small articulated oral spines ( Fig. 20A View Figure 20 ) ( Osburn, 1950 recorded 2–4 oral spines), except for the ancestrula which is tatiform and has numerous spines encircling the opesia. The ovicell is produced by a flat distal kenozooid ( Figs 20A- F View Figure 20 , 22E View Figure 22 ). The roof of the ovicell is constructed of 6–7 radial ribs which meet at the summit of the ovicell ( Fig. 20A- F View Figure 20 ). Ribs are joined to their neighbours via short lateral fusions, leaving rows of bean-shaped lacunae between fusions ( Figs 20D, E View Figure 20 , 22E View Figure 22 ). Sea water can apparently gain entry into the brooding cavity through these lacunae in living colonies. The two proximalmost ribs are significantly wider than the others and form a transverse bar that overlies the distal part of the maternal zooid ( Figs 20A, B View Figure 20 , 22E View Figure 22 ). The medial part of each rib is concave and possesses a surface fabric typical of cryptocysts, although without granulation. Therefore, a longitudinal strip of hypostegal coelom can be inferred to be present. The concave medial strips belonging to the most proximal pair of ribs are always joined, indicating continuity of the hypostegal coelom across the top of the ovicell. Similar strips in the more distal ribs may also reach the summit of the ovicell and join the strips of other ribs ( Fig. 20A, B View Figure 20 ). However, in other instances cryptocystal strips terminate before contacting those of neighbouring ribs, indicating that the hypostegal coeloms would not have linked with those of the other ribs.
Externally, each costa has a basal pore leading to a short canal through which the costal hypostegal coelom apparently communicates with the main visceral coelom of the kenozooid that forms the ooecium ( Figs 20D View Figure 20 , 22E View Figure 22 ). The entrance to this canal from the inside is identical in appearance to the pores leading to the pore-chambers ( Fig. 20E View Figure 20 ). Pores opening into costae and pore chambers alternate around the internal perimeter of the ovicell ( Fig. 20D View Figure 20 ). Unusually for ovicells, the frontal wall of the brooding kenozooid that forms the floor of the ovicell is membranous rather than being a calcified gymnocystal layer. The boundary between the main kenozooidal coelomic cavity and the brooding cavity of the ovicell is clearly seen as a demarcation line separating areas of different skeletal ultrastructure ( Fig. 20E, F View Figure 20 ). Ovicells are presumed to be cleithral.
Remark: Osburn (1950) was the first to describe Bellulopora bellula (as Colletosia ) from the Recent of the Gulf of California and the Pleistocene of southern California. He noted ‘the primary ovicell [possibly, kenozooid]’ that ‘is small..., smooth, hemispherical’, and ‘soon becomes covered by an ectooecium which is composed of 3 or 4 pairs of radiating costules’ (pp. 188– 189). Larwood (1962: 111) repeated Osburn’s description. However, Moyano (1984), when describing B. bellula from central Chile, came very close to understanding the unusual and enigmatic structure of its ovicell. He correctly recognized the kenozooidal nature of the ovicell in this species, and stated that ‘the ovicell external wall (= ectooecium) has the same structure as the frontal shield’ (p. 62). However, he thought that the hypostegal coelom of each costa connected with ‘a dietella [basal pore chamber] lying at the base of the lateral walls’ (p. 59; see also Moyano, 1991). He also found the ‘horizontal membrane that seems to be homologous to the frontal zooidal membrane’ (p. 62), but, unfortunately, did not appreciate that it represented the floor of the ovicell. Moyano described ‘a calcified globular structure (= the entooecium)’ positioned ‘inside [?] the ectooecium’. This is actually part of the peristome of the maternal zooid
A
B
C
D
E underlying the ovicell opening ( Figs 20C View Figure 20 , 22E View Figure 22 ) (for further discussion, see Gordon, 2000; Ostrovsky, 2002).
Brief descriptions of this species were also given by Lagaaij (1963) and Moyano & Melgarejo (1977), based on material, respectively, from the Gulf of Mexico and central Chile.
Additional data from the literature: There are several Recent cribrimorphs described in the literature whose ovicells resemble costate frontal shields and apparently have the same structure. Among these are Figularia clithridiata ( Waters, 1887) , F. tahitiensis Waters, 1929 , F. pulcherrima Tilbrook et al., 2001 , Puellina modica Bishop & Househam, 1987 (see Ostrovsky, 2002).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Leptocheilopora
| Ostrovsky, Andrew N. & Taylor, Paul D. 2005 |
BELLULOPORA
| LAGAAIJ 1963 |