
Macropora, MACGILLIVRAY, 1895
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00179.x |
|
persistent identifier |
https://treatment.plazi.org/id/6F0C243F-FF95-2577-FF4E-64670256FB4D |
|
treatment provided by |
Diego |
|
scientific name |
Macropora |
| status |
|
GENUS MACROPORA MACGILLIVRAY, 1895 View in CoL MACROPORA CRIBRILIFERA MAPLESTONE, 1901
( FIGS 14A- C View Figure 14 , 23B View Figure 23 , 24R View Figure 24 )
Material: NMV P311815. Lower Miocene, Mount Gambier Limestone, limestone quarry about 1 km west of Mount Shanck (37∞57¢S, 140∞43.2¢E), South Australia, collected by P. E. Bock.
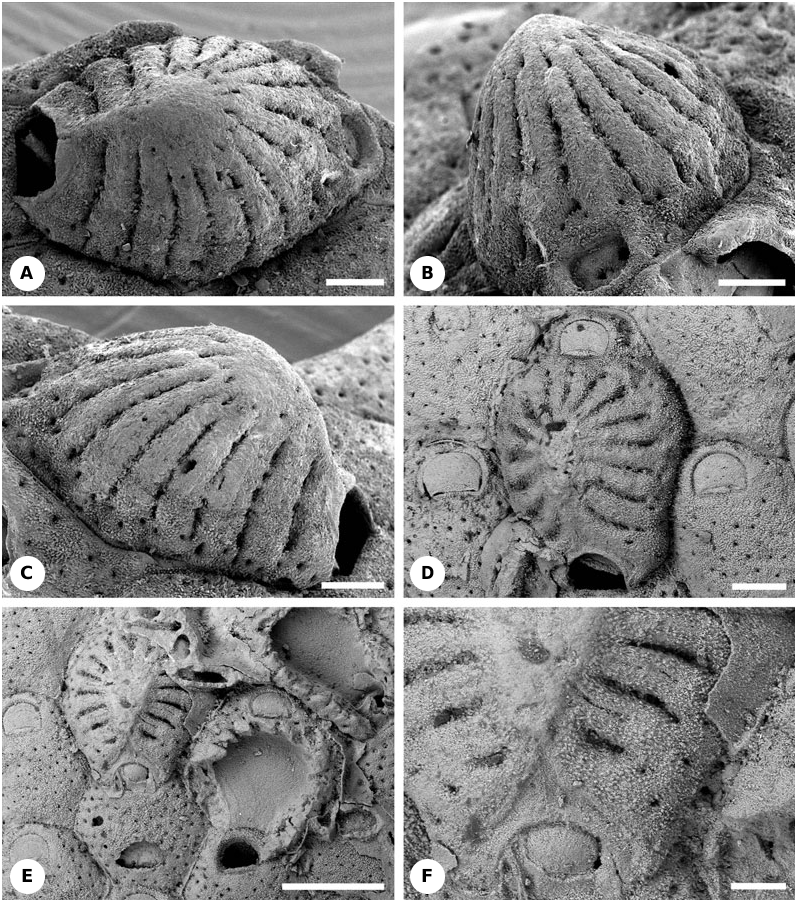
Description: The hyperstomial ovicells are normally constructed by distal autozooids but one observed example was formed by a distal kenozooid. Ovicells are large, occupying the entire zooidal frontal surface except for the orifice/opesia, and consist of 16 spinelike ribs (costae) arranged in a radial pattern and separated by distinct slits ( Figs 14A- C View Figure 14 , 23B View Figure 23 , 24R View Figure 24 ). The two most proximal ribs are wider than the others ( Fig. 14A, C View Figure 14 ), and the shortest ribs are sometimes found in the distal part of the ovicell ( Fig. 14B View Figure 14 ). The costae are flattened, with the lumen confluent with the chamber of the distal brooding zooid. Their ends are fused along the midline of the ovicell to form a median ridge. Laterally, the costae are also fused periodically with neighbouring costae. Intercostal slits have the appearance of rows of elongated pores ( Fig. 14A- C View Figure 14 ). Usually there are one or two pores on each side of the costal bases, with additional pores occasionally developed further along the costae ( Fig. 14A- C View Figure 14 ). Judging from the presence of pores and the granular external texture, each costa was enveloped (at least externally) by cryptocyst, and the ovicell roof in this fossil species was probably originally covered by an outer membrane with a hypostegal coelom beneath. This coelom would have been confluent both with the hypostegal coelom of the distal brooding zooid as well as the visceral coelom of the maternal zooid through the pores in the costal bases ( Fig. 23B View Figure 23 ). Since the majority of the frontal surface of the distal, brooding zooid is occupied by the ovicell, parietal muscles deforming the frontal membrane during tentacle crown protrusion presumably passed through the pores in the costal bases. The slightly concave ovicell floor is mainly gymnocystal, as are the undersides of the costae facing into the brooding cavity. The archlike opening of the ovicell is placed just above the maternal zooid orifice, probably being closed by the zooidal operculum (cleithral type).
Remarks: Maplestone (1901) described and illustrated Macropora cribrilifera from the Miocene of Australia (Mitchell River, near Bairnsdale, Victoria) (see also Canu & Bassler, 1929). SEM images of this species can be found on the website of P. E. Bock: (http:// www.civgeo.rmit.edu.au/bryozoa/cheilostomata/ macroporidae/macrcri.html). Superior preservation of the ovicells depicted here shows that each costa (except the most proximal pair) has a medial longitudinal keel. It is possible that this keel is a very narrow longitudinal strip of gymnocyst. The surfaces of both the median ridge of the ovicell as well as the perimeter around the opening of the ovicell are smooth, possibly also representing gymnocyst.
MACROPORA WAIMATUKUENSIS ( UTTLEY, 1949)
( FIGS 14D- F View Figure 14 , 24R View Figure 24 )
Material: CM zb51, holotype,? Miocene , right bank of Waimatuku River, Southland, New Zealand . NIWA, Eocene , Runangan (Priabonian), Alma , Oamaru, North Otago, New Zealand. NHM BZ5201 , Eocene, Runangan (Priabonian), Fortification Road cutting, Oamaru, North Otago, New Zealand, P. D. Taylor & M. J. Weedon Collection .
Description: The hyperstomial ovicell occupies the entire frontal surface of the distal brooding autozooid, apart from the orifice, and is constructed of 16 costae/ ribs arranged radially with slits between ( Figs 14D- F View Figure 14 , 24R View Figure 24 ). The costae are hollow with their lumens fused distally. Surface texture is granular, and there is an oval pore at the base of some of the costae. Thus, the costal surface is cryptocystal and would have been enveloped by hypostegal coelom with an outer membranous wall during life. Possible exceptions are the central area of the ovicell roof (medial ridge) where the costae are fused ( Fig. 14D, F View Figure 14 ), and the narrow stripes along the midline of some (all?) costae that are possibly gymnocystal. The arch-like opening of the ovicell is formed by the widest proximal costae. It is placed just above the maternal zooid orifice, and would have been closed by the zooidal operculum (cleithral type).
Remarks: Uttley (1949: 180) described and illustrated ovicells consisting of 16–18 ‘rounded costules’ in this species (as Macroporella ) from the Lower Oligocene (Whaingaroan) of Kakanui, North Otago, and the Lower Miocene of Waimatuku River, Southland, New Zealand. Additionally, he mentioned ‘costulate’ ovicells in M. waimatukuensis var. trisinuata Uttley, 1949 from the Upper Oligocene of North Otago and Middle Miocene (Waiauan) of North Canterbury.
( FIGS 15A, B View Figure 15 , 23B View Figure 23 , 24R View Figure 24 )
Material: NHM BZ5202–3 , Miocene, Otaian (Burdigalian), Waitemata Group, Parnell Grits, Faulkner Bay- Waikowhai section, Manukau Harbour, Auckland, New Zealand, P. D. Taylor Collection .
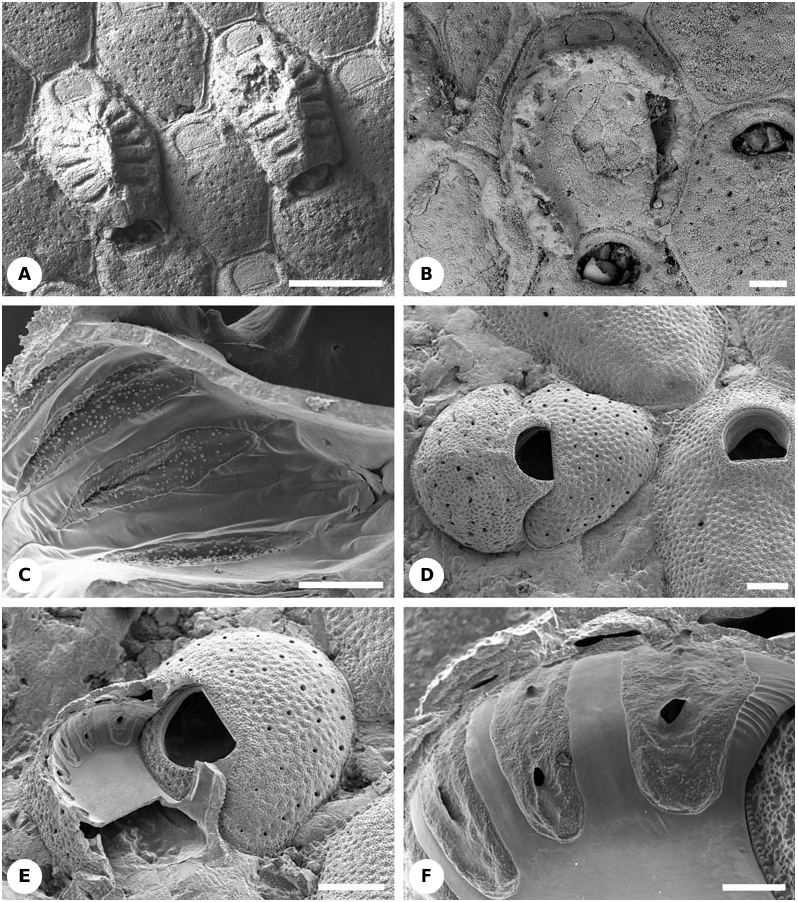
Description: The hyperstomial ovicell is very large, occupying the entire nonorificial frontal surface of the distal autozooid except for the orifice, and consists of 14–15 costae arranged in a radial pattern with broad intercostal slits ( Figs 15A View Figure 15 , 23B View Figure 23 , 24R View Figure 24 ). The two most proximal costae are wider than the others. A median ridge or boss is formed by the fusion of the distal ends of the flattened costae. The surface texture of the costae is granular, resembling the cryptocystal frontal shield. Therefore, it can be inferred that the costae were covered by a hypostegal coelom beneath a membranous external wall in the living bryozoan ( Fig. 23B View Figure 23 ). Worn ovicells expose the flat floor and show the presence of pores between the costal bases ( Fig. 15B View Figure 15 ). The arch-like opening of the ovicell is placed just above the maternal zooid orifice, and was probably closed by the zooidal operculum (cleithral type). Distal (brooding) autozooids have noticeably smaller orifices than nonbrooding autozooids.
( FIGS 15C View Figure 15 , 24R View Figure 24 )
Material: NIWA, Recent , NIWA Stations Kah 0204/22 (34∞4.68¢S, 174∞4.72¢E, depth 610 m) and Kah 0204/27 (34∞7.21¢S, 174∞5.64¢E, depth 554 m), Cavalli Seamounts, New Zealand, collected 16 April 2002 .
Description: Zooids possess 4–6 small, articulated oral spines. Brooding (distal) autozooids have large, hemispherical hyperstomial ovicells that occupy all of the zooid frontal surface except for the orifice. Maternal autozooids are shorter and wider than sterile autozooids. The opening of the ovicell is placed just above the orifice of the maternal zooid and is closed by the operculum (cleithral type). Externally, the calcified ovicell roof has a cryptocystal texture with pits, 12 short radial fissures and a medial furrow. The outer body wall above the ovicell calcified wall was evidently membranous, enclosing a hypostegal coelom confluent with that of the brooding zooid. Broken ovicells reveal that the internal structure of the roof comprises flattened, hollow costae, proximally with slits flanked by cryptocystal calcification ( Fig. 15C View Figure 15 ) but distally (at the summit of the roof) comprising fused, entirely gymnocystal costae. Costal lumens are confluent through slit-like openings with the perigastric cavity of the distal zooid but not with one another. The ovicell floor is gymnocystal.
MACROPORA UTTLEYI LÓPEZ DE LA CUADRA & GARCÍA GÓMEZ, 1997
( FIGS 15D- F View Figure 15 , 24R View Figure 24 )
Material: NIWA, Recent , NIWA Stations Kah 0204/22 (34∞4.68¢S, 174∞4.72¢E, depth 610 m) and Kah 0204/27 (34∞7.21¢S, 174∞5.64¢E, depth 554 m), Cavalli Seamounts, New Zealand, collected 16 April 2002 .
Description: Large hyperstomial ovicells are produced by a flat distal kenozooid that forms the base of the brood chamber. Maternal autozooids are smaller than the other autozooids ( Fig. 15D- F View Figure 15 ). The arch-like opening of the ovicell is placed just above the orifice of the maternal zooid, apparently being closed by the zooidal operculum (cleithral type). Externally, the calcified ovicell roof has a granular cryptocystal fabric with pits and pores arranged in an ill-defined radial pattern. It was evidently covered by an external membranous wall enclosing a hypostegal coelom. Broken ovicells show that the roof actually consists of about a dozen fused and flattened costae ( Figs 15E, F View Figure 15 , 24R View Figure 24 ). The surfaces of costae facing into the brooding cavity are gymnocystal, separated from one another by porous bands of cryptocyst. The pores connect the external ooecial hypostegal coelom with longitudinal hypostegal cavities between the costae on the ooecial internal surface. Coeloms within the lumens of the costae ( Fig. 15F View Figure 15 ) are apparently confluent with the perigastric cavity of the brooding zooid but not with each other. The ovicell floor is gymnocystal, its calcification smoothly continuous with that of the costae.
Remarks: López de la Cuadra & García-Gómez (1997) described and illustrated the radially arranged pores in the ovicells of this species from off Three Kings Island, New Zealand.
MACROPORA LEVINSENI BROWN, 1952 View in CoL
( FIGS 16A- C View Figure 16 , 17A, B View Figure 17 , 23E View Figure 23 , 24R View Figure 24 )
Material: NIWA, Recent, NIWA Station U1012, depth 35 m, east of New Zealand (39∞56.35¢S, 174∞25.55¢E).
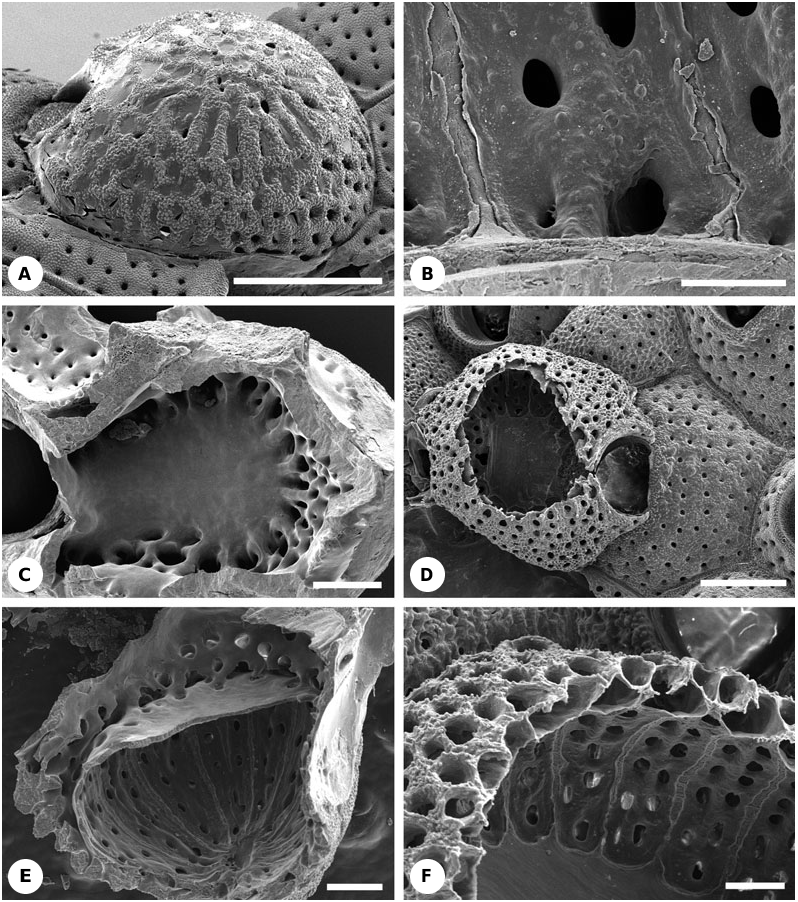
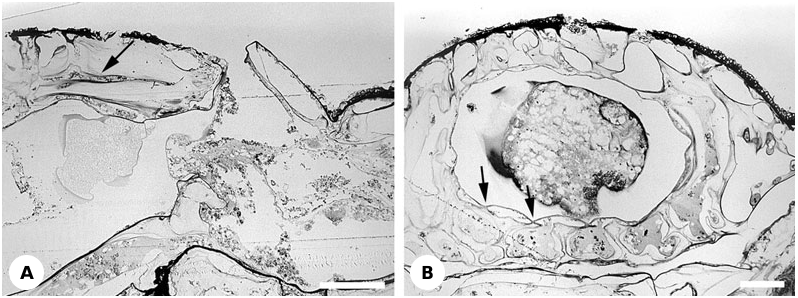
Description: The large hyperstomial cleithral ovicell is produced by the distal kenozooid ( Figs 16A, C View Figure 16 , 23E View Figure 23 ) which has pore chambers in contact with neighbouring autozooids. Compared with other autozooids, maternal autozooids are shorter whereas ovicells are longer. The calcified part of the ooecium consists of about 10–12 hollow costae embedded in a perforated cryptocystal matrix ( Figs 16B View Figure 16 , 17A View Figure 17 , 23E View Figure 23 , 24R View Figure 24 ). This complex wall is enveloped, both externally and internally ( Figs 16A View Figure 16 , 17A, B View Figure 17 , 23E View Figure 23 ), by the membranous outer body wall with its underlying hypostegal coelom. The external (upper) membranous ovicell wall is thicker than the internal (lower) one. Basal parts of the costae facing into the brooding cavity are gymnocystal ( Fig. 16B View Figure 16 ), not covered by the membranous wall and associated hypostegal coelom. In cleaned specimens longitudinal grooves visible on the internal ovicell surface correspond with costae.
From the outside the skeletal ooecial wall is a latticework of radial ribs, with granular surface texture, and subcircular and elliptical pores ( Fig. 16A View Figure 16 ). However, only some of these ribs correspond to the costae visible internally; others are exclusively cryptocystal. The subcircular pores are mainly concentrated around the ovicell base, and the elliptical pores on the ovicell roof. Pores lead to the anastomosing lacunar system piercing the cryptocystal matrix and connecting the upper and the lower hypostegal coeloms ( Figs 17B View Figure 17 , 23E View Figure 23 ). Costae, except for their basal parts facing into the brooding cavity, are covered with cryptocyst and their narrow coelomic cavities open into its lacunar system. In sections of decalcified specimens the costal skeleton stains differently from the surrounding cryptocyst. The arch-like opening of the ovicell is placed just above the orifice of the maternal zooid and is closed by the zooidal operculum (cleithral).
Most of the ovicell floor is gymnocystal except for the periphery where the costal bases and intervening cryptocystal overgrowths are covered by a membranous wall continuous with the internal membranous wall. Viewed from underneath, the peripheral area of the ovicell floor is also perforated by large and small pores ( Figs 16C View Figure 16 , 23E View Figure 23 ). Some of the large pores lead to the costal cavities, thereby connecting the costal coeloms with the kenozooidal coelom. Others are either fused with the former or pierce the floor linking the kenozooidal coelom with the hypostegal coelom above the ovicell floor.
Remarks: This extant species was first described as Macropora grandis (Hutton) var. levinseni by Brown (1952) based on material from Wanganui, New Zealand. There is at least one additional species, Macropora grandis (Hutton, 1873) , recorded from the Lower Miocene-Recent of Australia and New Zealand, with a similar or identical ovicell structure (for synonymy and localities, see Brown, 1952; Gordon, 1984; López de la Cuadra & García-Gómez, 1997).
Gordon (1984: 57) noted that the ovicell can be ‘resting on a distal [auto]zooid’ as well as on a distal kenozooid in Macropora grandis . Following Brown (1952), he also commented that there was no distal kenozooid in some instances, and ‘the ovicell apparently rests directly on the substratum’. Judging from the ovicell structure in M. levinseni , this statement is almost certainly incorrect and is based on the existence of ovicells formed by very low and flat kenozooids that are difficult to observe.
MACROPORA POLYMORPHA (PHILIPPS, 1899)
( FIGS 16D- F View Figure 16 , 24R View Figure 24 )
Material: NIWA, Recent, NIWA Station Z 9695, depth 89 m, Tom Bowling Bay , northern North Island, New Zealand (22∞00¢S, 173∞00.01¢W), collected 27 January 1999 .
Description: Zooids possess 3–4 small, articulated oral spines. The large hyperstomial ovicell is produced by a kenozooid ( Fig. 16D, E View Figure 16 ) distal of the maternal zooid which has pore chambers in contact with neighbouring autozooids. Compared to other zooids, the maternal autozooid is shorter whereas the ovicell is about the same size. Externally, the ooecial wall has a granular surface with oval or subcircular pores in radial rows ( Fig. 16D, F View Figure 16 ) and is clearly cryptocystal. The calcified part of the ooecium consists of more than a dozen hollow costae, embedded in a perforated cryptocystal matrix ( Figs 16E, F View Figure 16 , 24R View Figure 24 ). This wall is apparently covered by a membranous wall, with underlying hypostegal coelom, on both the external and internal sides. The underside of the ooecial roof facing into the brooding cavity has narrow, raised strips of smooth, nonporous skeleton separated by broader regions of porous obviously cryptocystal skeleton ( Fig. 1E, F View Figure 1 ). Each strip corresponds to a costa, having a short gymnocystal base. However, it is uncertain whether the narrow strips are gymnocystal; similar regions in M. levinseni are cryptocystal. Pores in the walls of the ooecium lead to the wide costal coelomic cavities and anastomosing lacunar system connecting the inferred upper and lower hypostegal coeloms ( Fig. 16F View Figure 16 ). The arch-like opening of the ovicell is placed just above the maternal zooid orifice, being closed by the zooidal operculum (cleithral).
Most of the ovicell floor is gymnocystal ( Fig. 16D View Figure 16 ). However, the peripheral region is cryptocystal and perforated by pores ( Fig. 16D, F View Figure 16 ). Some of the larger pores lead into the costal cavities whereas others possibly pierce the floor of the ovicell, thereby connecting the kenozoidal coelom with the hypostegal coelom above the ovicell floor.
Remarks: López de la Cuadra & García-Gómez (1997) described and illustrated ovicells with radially arranged pores in furrows in material of this species from the Loyalty Islands (Pacific Ocean).
Additional data from the literature: Uttley (1949: 178) described and illustrated ‘strongly costulate’ ovicells consisting of 18–28 ‘ribs’ in M. retusa ( Uttley, 1949) (as Macroporina ) from the Whaingaroan (Lower Oligocene) of Kakanui, North Otago, New Zealand. Scanning electron microscopy of Uttley’s material shows that the ovicell is formed by a distal kenozooid and constructed of 19 costae whose tips are fused along the median suture. Costal fusion leaves two groups of large oval slits, an upper group between the costal tips and a lower group between the costal bases. Each costa has a longitudinal furrow, possibly gymnocystal, whereas the rest of the surface has a cryptocystal relief. The base of the ooecium is encircled by round pores.
Radially striated ovicells with large marginal pores were described and figured by López de la Cuadra & García-Gómez (1997) in M. georgiensis López de la Cuadra & García Gómez, 1997 . In this species from the Recent of South Georgia (Atlantic Ocean) they illustrated developing ovicells, showing the gymnocystal ovicell floor, bases of the costae and intervening cryptocystal overgrowths with pores similar to those seen in M. levinseni , M. polymorpha (see above) and also in M. centralis MacGillivray, 1895 (Miocene, Muddy Creek, south-eastern Australia). The latter species has been depicted on the web-site of P. E. Bock: (http://www.civgeo.rmit.edu.au/bryozoa/cheilostomata/ macroporidae/macrcri.html). Ovicells with radial striations and marginal slits also occur in M. africana Hayward & Cook, 1983 (Recent, eastern South African coast), and in M. operculata ( Canu, 1908) (as Hippoporina ) (Miocene, Chubut, Patagonia, Argentina). These strongly suggest a similar or identical ovicell structure to the species described above, with costae overgrown by cryptocyst.
FAMILY CRIBRILINIDAE HINCKS, 1879


| CM |
Chongqing Museum |
| NIWA |
National Institute of Water and Atmospheric Research |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Macropora
| Ostrovsky, Andrew N. & Taylor, Paul D. 2005 |
MACROPORA LEVINSENI BROWN, 1952
| Brown, Recent 1952 |