
Araucoderus gloriosus ( Alexander, 1920 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4158.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:3FA278A1-1407-47BE-9A79-DEFF9A03B3E4 |
|
DOI |
https://doi.org/10.5281/zenodo.5668879 |
|
persistent identifier |
https://treatment.plazi.org/id/6D35EC12-0623-FF81-A3F0-3FA6E603FCC1 |
|
treatment provided by |
Plazi |
|
scientific name |
Araucoderus gloriosus ( Alexander, 1920 ) |
| status |
|
Araucoderus gloriosus ( Alexander, 1920) View in CoL
Tanyderus gloriosus Alexander, 1920: 13 View in CoL : 5.
Radinoderus gloriosus Alexander, 1927: 189 : 9. Araucoderus gloriosus Alexander, 1929: 1 View in CoL : 228.
Diagnosis. Egg: The only Tanyderidae known in this stage. Larva: cuticle translucent, varying from light grey to tan in color; head orange to brown in color; mandible with three prominent teeth; endocarina (glandular duct) present; postmentum without teeth, flat to convex anteriorly. Pupa: Body elongate, cylindrical; head with tubercles, antennal sheath lying between wing margin and midleg; respiratory organs inconspicuous, undivided and tubular; abdominal segment VIII with digitiform lobes inserted posterolaterally, posterior to spiracle. Male: Cuticle with variegated coloration, light- to dark brown; vestiture light brown; abdominal tergites dark brown, with irregular, pale markings anteriorly on either side of median; wing more than 14 mm in length, infuscated, with pigment arranged in transverse bands; supernumerary cross-veins absent; male terminalia: with varying degree of rotation, usually 90 degrees, involving segments 7–9; dorsal bridge of paramere with pair of projections anterolaterally; lateromedial element of paramere with small convex anterior lobe bearing 4–6 setiform sensilla, with a dentate mesal lobe set with five dissimilar projections, with a fleshy convex lobe inserted proximally, set with many small trichoid sensilla; dorsomedial element “I-shaped”, articulated basally with dorsal bridge, articulated apically with lateral ejaculatory processes of aedeagus; parameral lobe at gonocoxite base reniform in lateral view, with several prominent setiform sensilla inserted subapically; aedeagus trifurcate, with point of trifurcation asymmetrical in dorsal view in some specimens; phallotrema flared slightly. Female: coloration, head, thorax, legs and wings as in male; spermatheca pyriform; ducts uniformly sclerotized, annulated and unpigmented.
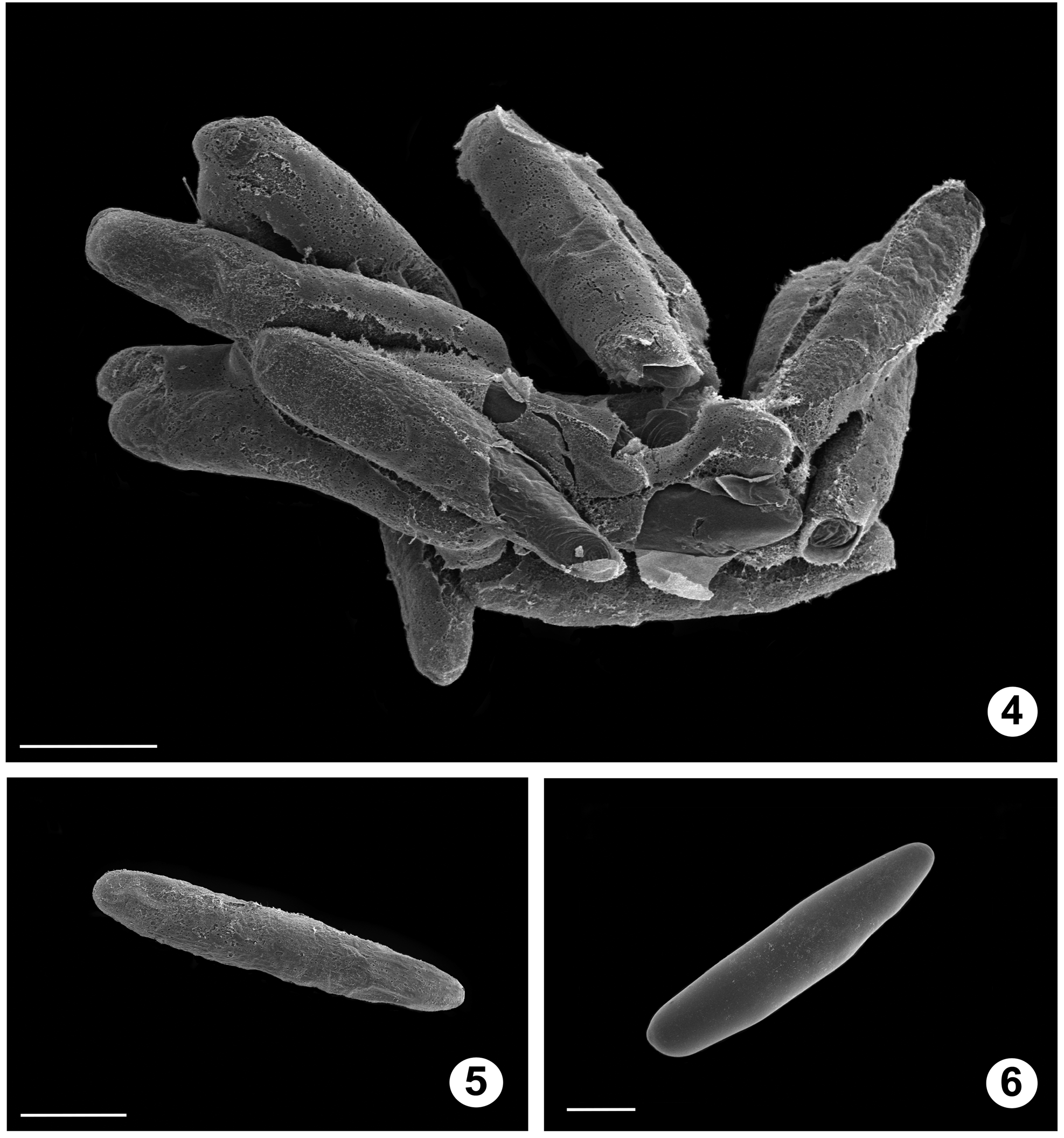
Description. See also Alexander (1920). Egg ( Figs. 4–6 View FIGURES 4 − 6 ): elongate-ovoid, white, lacking microsculpture; micropile not evident; laid in clusters of 10 or more.
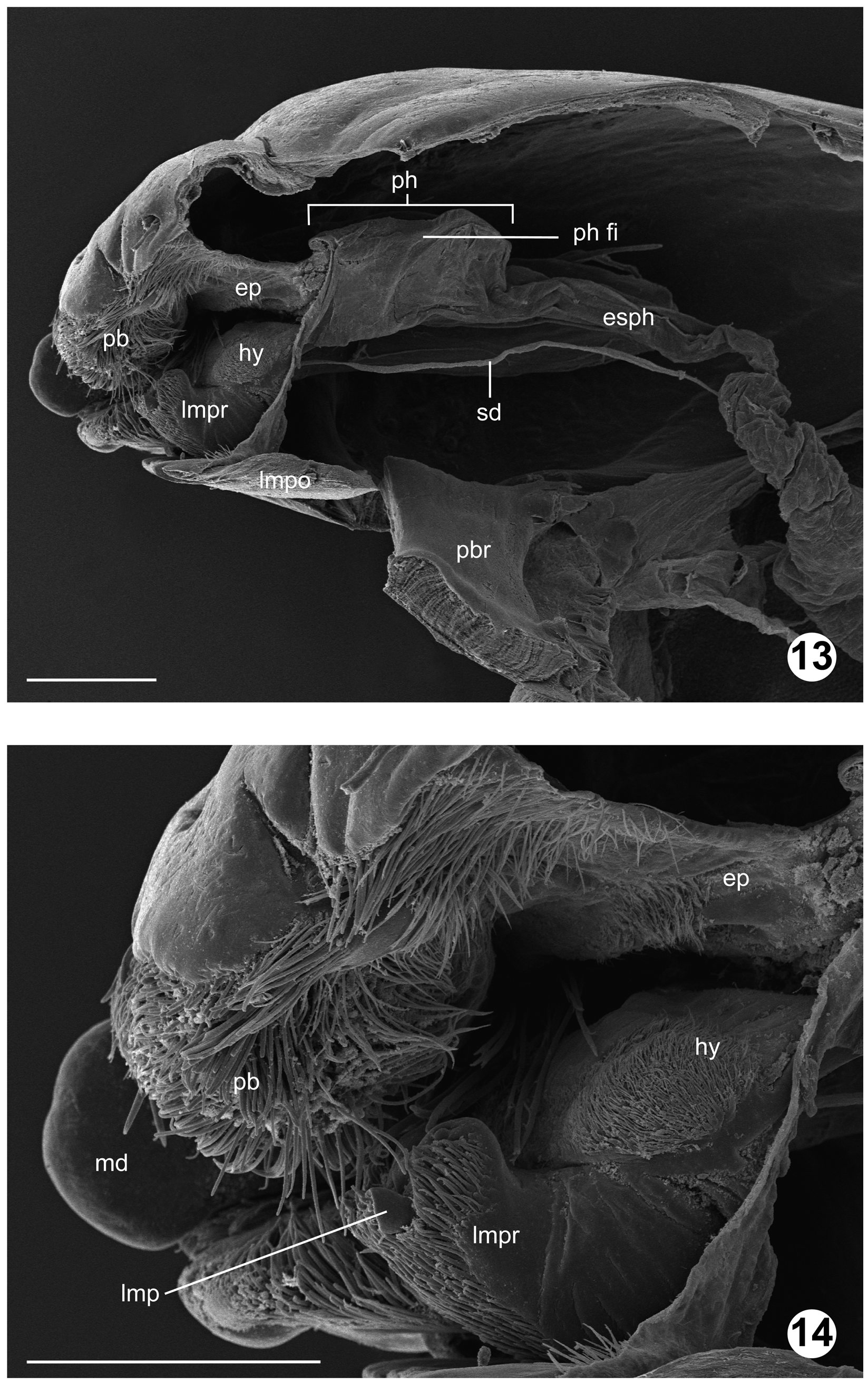
Larva IV instar ( Figs. 7–21 View FIGURES 7 − 10 View FIGURES 11 − 12 View FIGURES 13 − 14 View FIGURES 15 − 21 , 49, 50, 52, 53 View FIGURES 46 – 53 ): Measurements (n = 3), total length 31.34 mm (30.43–32.20), head width 1.35 mm (1.33–1.43), head length 1.67 mm (1.58–1.80). Head capsule ovate, heavily sclerotized anteriorly, dorsoventrally compressed, rounded posteriorly, partially retracted into prothorax; stemmata not apparent; primordial adult eye well-developed; frontoclypeal apotome (frcly ap; Figs. 11 View FIGURES 11 − 12 , 16 View FIGURES 15 − 21 ) somewhat pentagonal in shape, enclosed by frontal sutures; coronal suture visible medially on posterior 1/3 of the head; mesal endocarina (ec; Fig. 11 View FIGURES 11 − 12 , 16 View FIGURES 15 − 21 ) ending medially on anterior clypeal margin; anterior clypeal margin (acm; Fig. 11 View FIGURES 11 − 12 , 16 View FIGURES 15 − 21 ) composed of two pairs of undifferentiated lobes, medial pair elongate-transverse, lateral pair bulbous; labrum as broad as medial endocarinal lobes combined, broad, fleshy, somewhat dorsoventrally compressed; paraclypeal setae paired (s 3, s 4; Fig. 11 View FIGURES 11 − 12 ); labral sclerite with paired setiform sensilla widely separated (s 2; Fig. 11 View FIGURES 11 − 12 ); palatal brush (pb; Figs. 12 View FIGURES 11 − 12 – 14,18) with paired setiform sensilla inserted dorsally (s 1; Fig. 11 View FIGURES 11 − 12 ); antenna (ant; Figs. 10 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 18, 20 View FIGURES 15 − 21 ) inserted anterolaterally; base partially shrouded by epicondyle (econ; Figs. 11 View FIGURES 11 − 12 , 16, 20 View FIGURES 15 − 21 ); two-segmented, basal segment more than twice the length of distal segment; apical segment with at least six inconspicuous, two conspicuous sensilla basiconica inserted apically (aps; Fig. 10 View FIGURES 7 − 10 ); mandibles (md; Figs. 7 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 14 View FIGURES 13 − 14 , 16, 18, 20 View FIGURES 15 − 21 ) well developed, symmetrical, pyramidal in shape, scoop-like, with three dentations apically; middle dentation projecting beyond others; oral surface of mandible with well-developed prosthecal brush dorsally, two setiform sensilla ventrally, adoral surface with two sensilla basiconica (s 12, s 13; Fig. 12 View FIGURES 11 − 12 ), anterior condyle well developed; maxillary stipes with one prominent digitiform sensilla anterodorsally (s 21; Fig. 7 View FIGURES 7 − 10 ), with cluster of microtrichia posteroventrally; galea globular (mxg; Figs. 7, 8 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 18 View FIGURES 15 − 21 ), partially sclerotized, with three apical setiform sensilla (s 18, s 19, s 20; Figs. 7, 8 View FIGURES 7 − 10 ); lacinia elongate (mxl; Figs. 7, 8 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 18 View FIGURES 15 − 21 ), covered with microtrichia adorally, bearing one seta (s 22; broken in Fig. 9 View FIGURES 7 − 10 ) anterolaterally; maxillary palpus (mxp; Figs. 7–9 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 18, 20 View FIGURES 15 − 21 ) unsegmented, cylindrical, slightly longer than wide, set with many sensilla basiconica apically, with apical sensilla globose to digitiform, arranged in whirls; cardo (mxc; Figs. 7, 8 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 18 View FIGURES 15 − 21 ) with three setiform sensilla (s 24–s 26; Fig. 8 View FIGURES 7 − 10 ), separated from stipes (mxs; Figs. 8 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 18 View FIGURES 15 − 21 ) by cardostipital ridge; postmentum (lmpo; Figs. 8 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 , 13 View FIGURES 13 − 14 , 18 View FIGURES 15 − 21 ) heavily sclerotized, quadrate basally, with anterior margin straight or slightly emarginated, concave; postgenal bridge (pbr; Fig. 13 View FIGURES 13 − 14 ) complete; prementum (lmpr; Figs. 7 View FIGURES 7 − 10 , 12 View FIGURES 11 − 12 –14,18) with saddle-shaped sclerite apically; labial palpus (lmp; Figs. 7 View FIGURES 7 − 10 , 14 View FIGURES 13 − 14 ) cylindrical, sclerotized, with six digitiform sensilla apically, two almost twice as long as the rest; glossae inconspicuous, (lmg; Fig. 7 View FIGURES 7 − 10 ) conical; hypopharynx (hy; Figs. 7 View FIGURES 7 − 10 , 13, 14 View FIGURES 13 − 14 ) cushion-like, with semicircular sclerite dorsally; pharyngeal filter (ph fi; Fig. 13 View FIGURES 13 − 14 ), housed within pharynx (ph; Fig. 13 View FIGURES 13 − 14 ) well developed, supported by two narrow, sclerotized oral arms originating from hypopharynx. Thoracic segments (pro, mes, met; Fig. 15 View FIGURES 15 − 21 ) equal in length, each bearing ten transverse, equally spaced 1–4 branched setae dorsally, four 2–4 branched setae laterally and six clusters of 1–3 branched setae ventrally; prothorax bearing pair of well-developed spiracles laterally (a spir; Fig. 15 View FIGURES 15 − 21 ), pro- and mesothorax slightly differentiated by tenuous intersegmental constriction; metathorax with circular and pad-like lobes ventrally. Abdominal segments (aI–aVIII & aA; Fig. 15 View FIGURES 15 − 21 ) with distinct intersegmental constrictions; abdominal segments I–IV with circular lobes ventrally, two simple setae, one bifurcated seta posterolaterally; segment VIII with one pair of posterior spiracles (p spir; Figs. 15, 17, 21 View FIGURES 15 − 21 ), one pair of elongate, annulated filaments extending beyond anal division; filaments with evenly spaced simple setae posteriorly; anal division (aA; Figs. 15, 17, 19, 21 View FIGURES 15 − 21 ) with one pair of elongate, annulated filaments dorsally, one pair of prolegs (pran; Figs. 19, 21 View FIGURES 15 − 21 ) ventrally; prolegs each with elongate, annulated filament inserted dorsoapically, with two rows of retractile hooks apically, basal row with 11–13 hooks, apical row with 12–13 slightly longer hooks; anal papillae (ap; Figs. 15, 17, 19, 21 View FIGURES 15 − 21 ) elongate-digitiform, twice the length of extended prolegs.
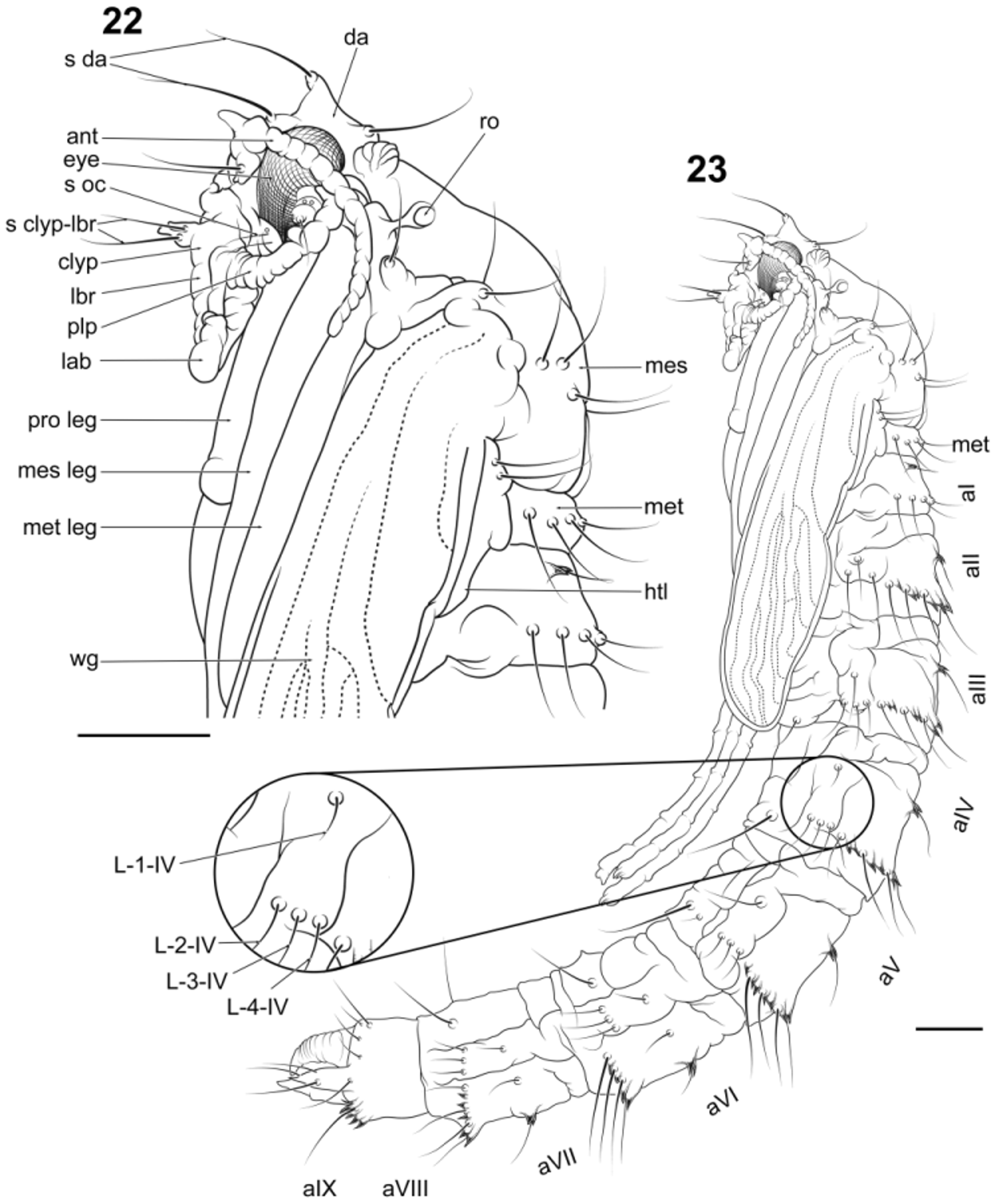
Pupa: Male: Measurements (n = 1), total length 18.40 mm, respiratory organ 0.50 mm (ro; Figs. 23 View FIGURES 22 − 23 , 25 View FIGURES 24 − 25 ). Body ( Figs. 22–25 View FIGURES 22 − 23 View FIGURES 24 − 25 , 51 View FIGURES 46 – 53 ) elongate, cylindrical, with cephalothorax circumference slightly larger than abdomen; mouthparts and thoracic appendages fused to each other and to remainder of cephalothorax. Head with tubercles; dorsal apotome (da; Figs. 23 View FIGURES 22 − 23 , 25 View FIGURES 24 − 25 ) apically with single sensilla (s da; Figs. 22 View FIGURES 22 − 23 , 24 View FIGURES 24 − 25 ), apotome ventrally abutting anterior margin of face; cephalic sclerite with one seta, sclerite fused anterolaterally to orbital portion of head capsule; antenna (ant; Figs. 22 View FIGURES 22 − 23 , 24 View FIGURES 24 − 25 ) extending posteriorly to between wing margin and midleg; clypeal apotome with pair of sensilla inserted basally; mouthparts restricted to area anterior of forecoxae; apex of labrum (lbr; Figs. 22 View FIGURES 22 − 23 , 24 View FIGURES 24 − 25 ) truncate; mandible present, maxilla absent; palpus (plp; Figs. 22 View FIGURES 22 − 23 , 24 View FIGURES 24 − 25 ) directed obliquely anterolaterally; labella divided medially; sensilla: setae arising from lobes; dorsal apotomals bilobed. Thorax: respiratory organ tubular, with numerous annulations, pores concentrated at apex; legs (pro leg, mes leg, met leg; Figs. 22 View FIGURES 22 − 23 , 24 View FIGURES 24 − 25 ) directed posteriorly, with tarsi extending posteriorly beyond wing apex, lying parallel to each other; metanotum (met; Fig. 22 View FIGURES 22 − 23 ) undivided medially, haltere (htl; Fig. 22 View FIGURES 22 − 23 ) extended posterolaterally before anterolateral margin of abdominal tergite 2 (aII; Figs. 23 View FIGURES 22 − 23 , 25 View FIGURES 24 − 25 ). Abdomen: circular in cross-section; tergites I–VIII with setae arranged in transverse rows on sclerotized bands bearing small coniform tubercles; pleurites II–VII with setae arranged in one caudal longitudinal row of three setae and one seta inserted in medial tubercle, adjacent to spiracle; sternites I–VIII with one pair of setae inserted posteriorly; segment VIII with posterolateral fleshy lobes bearing two setae posteriorly to spiracle; Segment IX (aIX; Figs. 23 View FIGURES 22 − 23 , 25 View FIGURES 24 − 25 ) without setae, a pair of posterior dorsomedial fleshy lobes present; terminal process apically tapered in both sexes. Female: Measurements (n = 1), total length 23.52 mm, respiratory organ 0.57 mm. In other details, same as male.
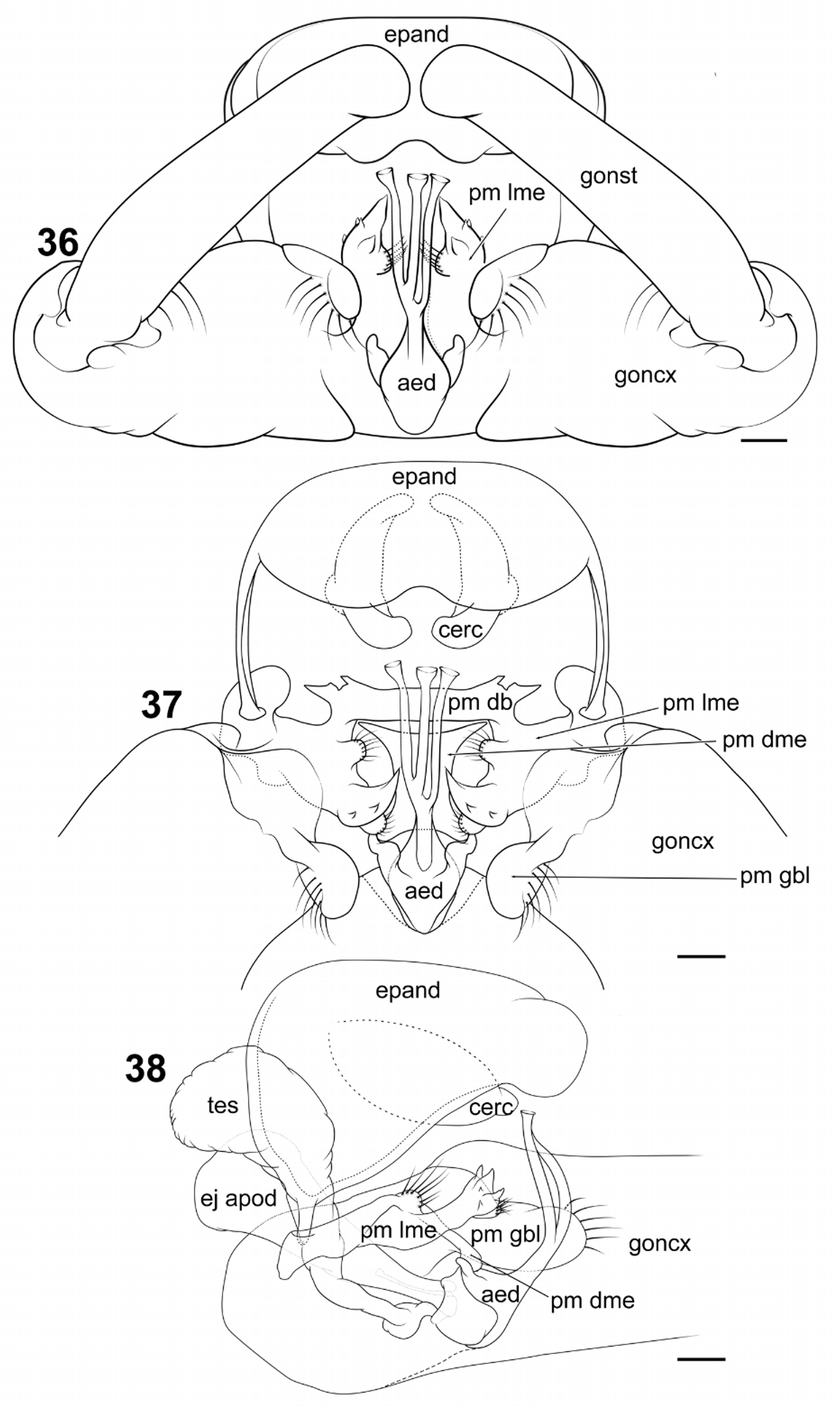
Male ( Figs. 1 View FIGURE 1 , 26–29 View FIGURES 26 − 29 , 32–38 View FIGURES 32 − 35 View FIGURES 36 − 38 , 46 View FIGURES 46 – 53 ): Measurements (n = 5), total length 14.66 mm (12.48–17.28); head length 1.48 mm (1.42–1.56); head width 1.22 mm (1.16–1.30); palpus length 1.54 mm (segment length average 0.15– 0.24–0.57–0.31–0.32); clypeus length 0.41 mm (0.40–0.44); clypeus width 0.37 mm (0.35–0.39); wing length 15.82 mm (14.14–18.08); wing width 3.75 mm (3.35–4.05). Head with mandible and maxilla poorly developed (md, mx; Fig. 27 View FIGURES 26 − 29 ); antenna with scape and pedicel glabrous, each with several setiform sensilla inserted distally; scape cylindrical (scp; Figs. 27, 28 View FIGURES 26 − 29 ), slightly larger than pedicel; pedicel spherical (ped; Figs. 27, 28 View FIGURES 26 − 29 ); flagellomeres numbering 15, all elongate, more than 3 times longer than wide, cylindrical; f 1 slightly longer than f 2; f 3–14 each with 4–5 trichoid sensilla inserted near base of flagellomere, arranged concentrically, as long or longer than flagellomere; f 1–15 each with several basiconic sensilla throughout; f 15 with 3 trichoid sensilla inserted apically. Cervical sclerites more than two times longer than wide. Wing ( Figs. 30, 31 View FIGURES 30 − 31 ) held away from body at angle in live resting individual; infuscate, with three transverse bands of pigment, basal-most band extending from h vein to posterior wing margin and slightly beyond proximal area of anal loop, medial band extending from midlength of Sc vein to posterior edge of C vein at distal end of anal lobe, passing through r-m and m-cu; apical-most band touching C vein on both sides of wing, appearing as an “X” from anterĺor margĺn of wĺng to apĺces of R4, M Z; costal cell yellow in color; cell r1 with two isolated, irregular, infuscated spots, one midlength of vein Rs and a second spot at cell apice; other spots, when present, variable among individuals. Halteres pale yellow at base, dark at apex. Legs light to dark brown, darker at both ends: Tibial spurs 0–2–2, symmetrical. Abdomen: segment I less than half as long as next segment, causing abdomen to droop ventrally; tergites dark brown with large irregular pale yellow markings basally on either side of medial line, each with dark yellow ovoid pattern centrally, basally. Terminalia ( Figs. 32–38 View FIGURES 32 − 35 View FIGURES 36 − 38 ) with torsion of segments 7–9 variable from 0 to 180 degrees; gonocoxites (goncx; Figs. 32, 33 View FIGURES 32 − 35 , 36– 38 View FIGURES 36 − 38 ) narrowly contiguous at base ( Fig. 33 View FIGURES 32 − 35 ), divergent from each other at origin, each nearly cylindrical, tapering slightly toward apex; setiform sensilla or setae alveoli distributed over dorsal, ventral and lateral surface, absent mesally; gonostylus (gonst; Figs. 32, 33 View FIGURES 32 − 35 , 36 View FIGURES 36 − 38 ) cylindrical, about 2/3 length of gonocoxite, slightly tapering at apex, curved medially, with prominent setae alveoli evenly distributed over surface, hirsute apically; hypandrium not evident; epandrium (epand; Figs. 32 View FIGURES 32 − 35 , 36–38 View FIGURES 36 − 38 ) slightly wider than long, with setae alveoli distributed over entire surface; proctiger recessed anteriorly, nested within epandrium; cercus (cerc; Figs. 37, 38 View FIGURES 36 − 38 ) inconspicuous, unmodified, setose; paramere subdivided into dorsal bridge (pm db; Fig. 37 View FIGURES 36 − 38 ), lateromedial element (pm lme; Figs. 36–38 View FIGURES 36 − 38 ), dorsomedial element (pm dme; Figs. 37, 38 View FIGURES 36 − 38 ) and parameral lobe at gonocoxite base (pm gbl; Figs. 32, 33 View FIGURES 32 − 35 , 37, 38 View FIGURES 36 − 38 ); dorsal bridge arch-shaped, with a pair of small projections anterolaterally; lateromedial element broadly interconnected with gonocoxite anteromesally, divided into three prominent lobes, anterior lobe small, convex, with 4–6 setiform sensilla, mesal lobe dentate, with five dissimilar projections, with fleshy convex lobe proximally bearing many small trichoid sensilla; dorsomedial element “I-shaped”, articulated basally with dorsal bridge, articulated apically with lateral ejaculatory processes of aedeagus; parameral lobe at gonocoxite base reniform in lateral view, with several prominent setiform sensilla inserted subapically, many smaller trichoid sensilla inserted on entire surface; ejaculatory apodeme (ej apod; Figs. 32–35 View FIGURES 32 − 35 , 38 View FIGURES 36 − 38 ) extending anteriorly to abdominal segment 7 terminus, laterally compressed, clavate at base, with surface corrugated; sperm sac (spm sac; Figs. 34, 35 View FIGURES 32 − 35 ) balloonlike, surrounded by aedeagus posteriorly, attached to ejaculatory apodeme anteriorly; aedeagus (aed; Figs. 32–38 View FIGURES 32 − 35 View FIGURES 36 − 38 ) narrowed abruptly at base, trifurcated at midlength; point of trifurcation asymmetrical in dorsoventral view; phallotrema flared slightly, placed between cerci when at rest; testes (tes; Figs. 32–35 View FIGURES 32 − 35 , 38 View FIGURES 36 − 38 ) pyriform, rounded anteriorly, differentiated from vasa deferentia only by width, shape; vasa deferentia (v d; Figs. 34, 35 View FIGURES 32 − 35 ), testes rugose in appearance; ejaculatory duct (ej dt; Figs. 34, 35 View FIGURES 32 − 35 ) lightly sclerotized, entering sperm sac through ejaculatory apodeme foramen.
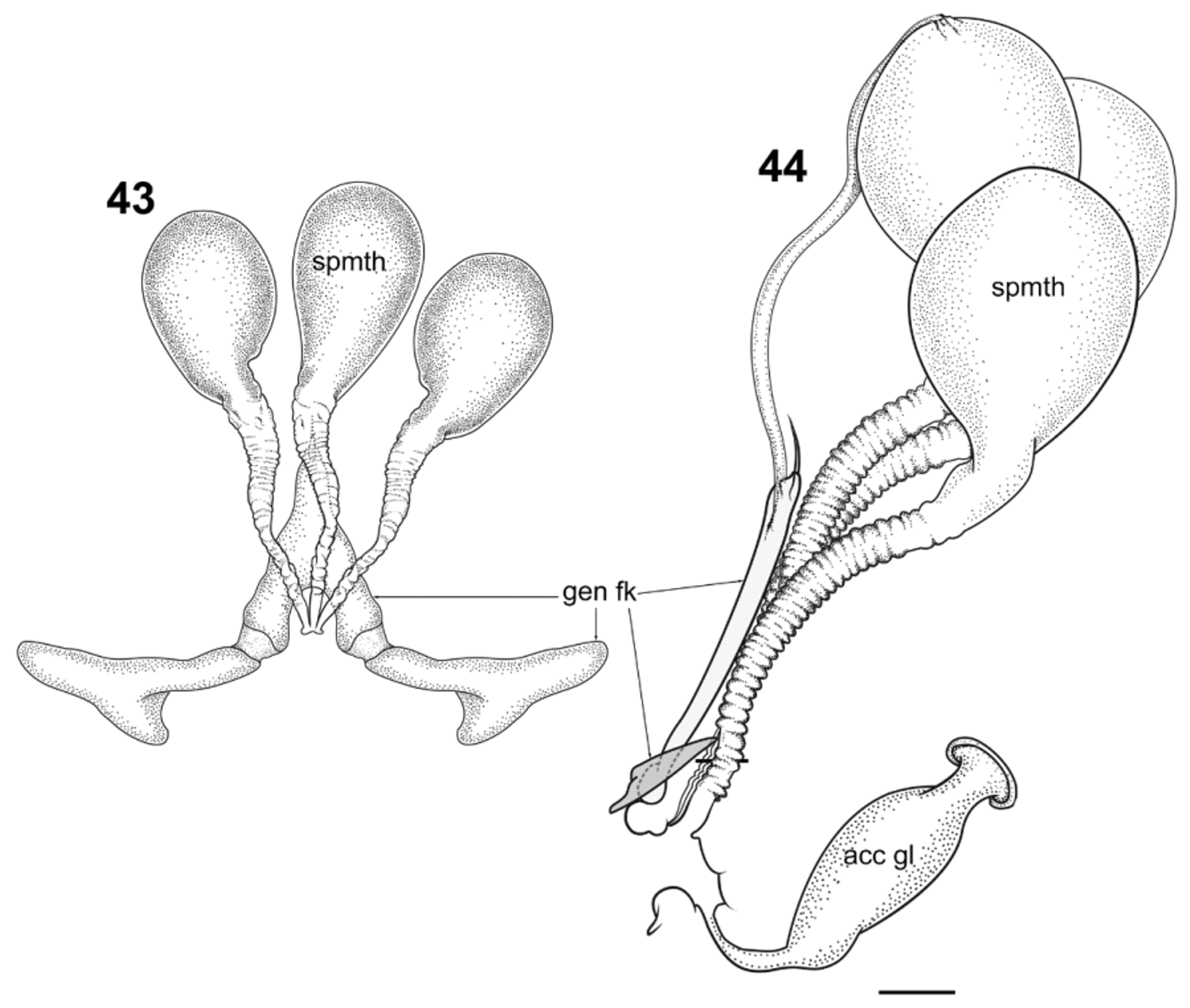
Female ( Figs. 39–45 View FIGURES 39 − 42 View FIGURES 43 − 44 View FIGURE 45 ): Measurements (n = 3), total length 21.02 mm (19.20–24.48); head length 1.51 mm (1.25–1.72); head width 1.23 mm (1.14–1.38); clypeus length 0.47 (0.37–0.55); clypeus width 0.40 (0.35–0.47); palpus length 1.80mm (palpomere length average 0.11–0.34–0.66–0.32–0.36); wing length 18.56 mm (16.15– 22.08); wing width 4.92 mm (4.00–5.96). Head with eyes as in male; mandible and maxilla twice as long as wide; antenna as in male. Wing as in male. Terminalia with posterior margin of sternite VIII broadly bilobate, strongly emarginate medially, emargination with “square U” shape; genital fork (gen fk; Figs. 40, 42 View FIGURES 39 − 42 , 43, 44 View FIGURES 43 − 44 ) transversely elongated posteriorly, more than eight times longer than wide, “Y”-shaped medially, acuminate anteriorly; spermatheca numbering three; corpora ovoid, with visible necks; spermathecal ducts annulated and uniformly sclerotized, more than twice as long as corpora, unpigmented.
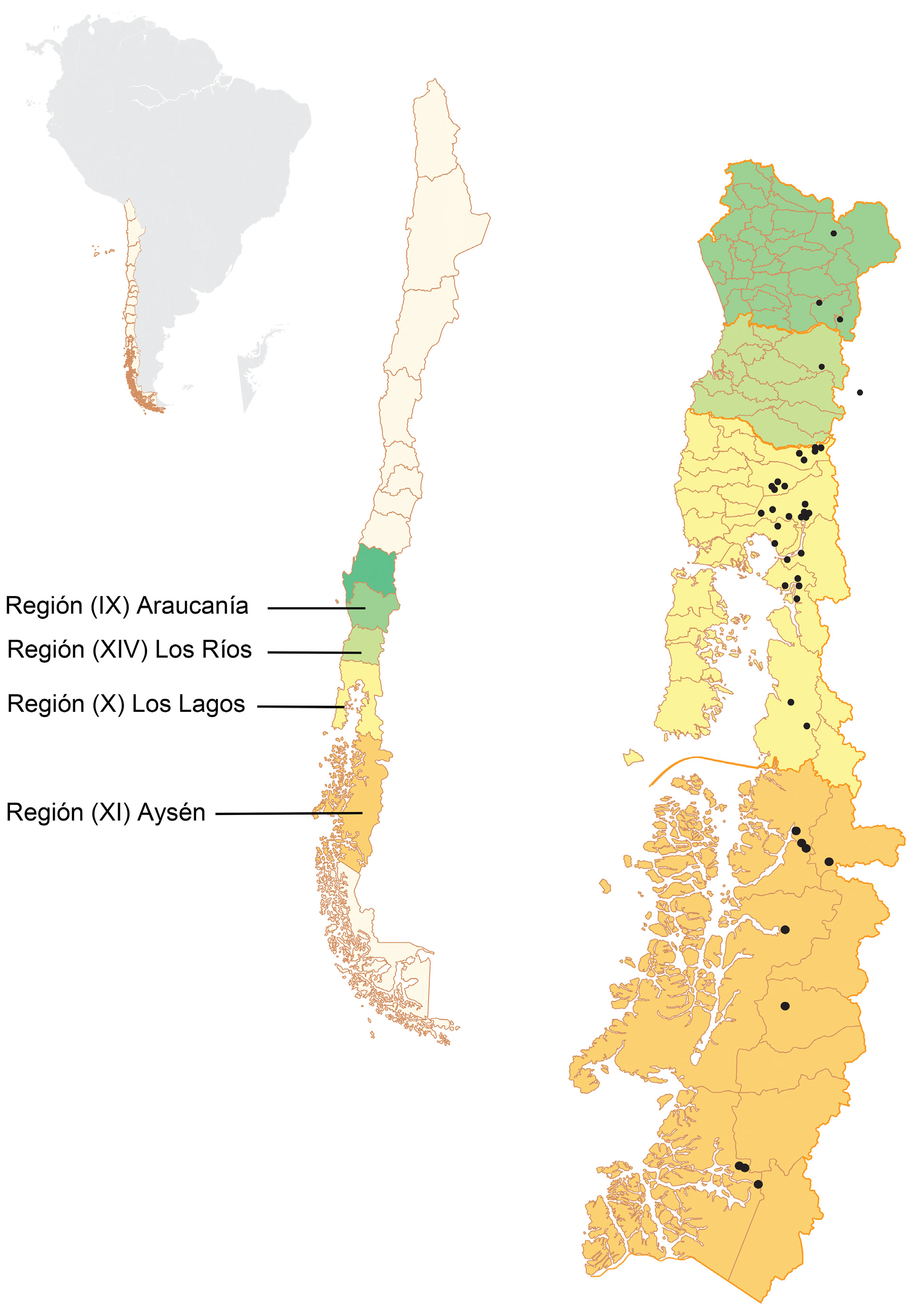
Distribution. Araucoderus is restricted to Patagonia. All confirmed records lie between latitudes 38°31’S and 48°02’S, ranging in altitude from 2 to 1700masl. A survey of the central regions of Chile yielded no specimens. There is one published Araucoderus record from Argentina ( Alexander, 1959).
Type material. Chile. Región XI (Aysén): Holotype Ƌ: Chile Rio Aysén, approximately, coll. P. Dusen, no date given; the specimen is deposited in two museums. Naturhistoriska Riksmuseet Stockholm , Sweden ( NHRM) and the USNM. The latter possesses only the wing, antenna and middle leg. Only the USNM material was examined during the current study.
Material examined. See Figure 2. Chile. Región IX (Araucanía): Estero el Rosario @ confluence with Estero la Cascada 39°23’12.93”S 71°41’46.27”W, 710masl, 3.x.2007, coll. G.W. Courtney, [2L]; Villarica N.R GoogleMaps . Río Quelhue @ northeast side Lago Villarrica 39°14.134’S 71°57.758’W, 230masl, 29.xi.2013, coll. R.I. Madriz, on riparian vegetation [1Ƌ] GoogleMaps ; Río Blanco Sur, downstream from Conguillío Road 38°31’16.62”S 71°50’17.70”W, 601masl, 19.xii.2013, coll. G.R. Curler, riparian vegetation [1♀]. Región XIV (Los Ríos): Estero Truful @ Hwy 203 crossing 39°49’29.97”S 71°59’53.63”W, 230masl, 4.xii.2013, coll. R.I. Madriz [3Ƌ]. Región X (Los Lagos): Casa Pangue Llanquihue, 4.xii.1926, coll. E.S. Shannon [1Ƌ]; Ensenada, Llanquihue, 2.i.1937, coll. E.C. Reed [1Ƌ]; Llanquihue Hornohuinco, xii.1968, coll. L.E. Pena [1Ƌ]; Lago Puyehue southeast shore, 10.ii.1978, coll. W.N. Mathis [1Ƌ] GoogleMaps ; Osorno Prov. Laguna el Espejo, 7.ii.1978, col. W.N. Mathis [1Ƌ]; Palena Río Amarillo ca. 28km southeast Chaitén, 23.i.1987, coll. C.M. & O.S. Flint, Jr., [1Ƌ] ; Llanquihue Prov. Río del Este @ Río Cochamó , 5km east Cochamó 41°30’S 72°16’W, 2–4.ii.1998, coll. N.E. Woodley [9Ƌ]; Prov. Llanquihue, {Cayutue} near Ralún 41°14’S 72°16’W, 12.ii.1998, coll. N.E. Woodley [2Ƌ]; Puyehue N.P., Río Anticura above Salto Río Anticura GoogleMaps , 40°40’16.14”S 72°10’12.88”W, 395masl, 4.xii.2008 coll. G.W. Courtney [1♂]; Puyehue N.P., Río Anticura above Salto Río Anticura GoogleMaps , 40°40’16.14”S 72°10’12.88”W, 395masl, 6.xii.2013, coll. R.I. Madriz [1Ƌ]; Puyehue N.P., Sendero Pudú 40°40’15.1”S 72°10’11.6”W, 393masl, 9.xii.2013, coll. S. Marshall & G.W. Courtney [1Ƌ]; Río Blanco @ Puente Río Blanco Arenales 41°03’04.6”S 72°39’46.8”W, 69masl, 9& 13.xii.2013, coll. R.I. Madriz & G.W. Courtney [2L, 1P, 2Ƌ, 1♀]; Puyehue N.P., Río Colorado @ Puente Arauco Sendero de Chile 40°40’15.5”S 72°08’43.9”W, 467masl, 10.xii.2013, coll. R.I. Madriz, G.W. Courtney & G.R. Curler [10L, 1P, 13Ƌ, 1♀] GoogleMaps ; Small creek ≈ 7km above Ralún on road to Cayetué 41°19’50.7”S 72°16’30.1”W, 240masl, 12.xii.2013, coll. R.I. Madriz & G.W. Courtney [6L, 5 Ƌ]; Cayetué Road, small creek ≈ 5.5k above Ralún 41°20'20.89"S 72°16'23.40"W, 150masl, 13.xii.2013, coll. G.W. Courtney, riffle kick sample [5L] GoogleMaps ; Río Pedregoso @ Puente Pedregoso 41°20’12.9”S 72°24’27.3”W, 55masl, 13.xii.2013, coll. R.I. Madriz [3Ƌ] GoogleMaps ; Río Blanco @ Puente Río Blanco , 40°58’36.5”S 72°35’01.9”W, 242masl, 14.xii.2013, coll. R.I. Madriz [2Ƌ] GoogleMaps ; Río Blanco @ Puente Río Blanco 41°00’30.2”S 72°34’13.9”W, 330masl, 14.xii.2013, coll. R.I. Madriz [2Ƌ] GoogleMaps ; Río Llaquepe @ Puente Llaquepe 41°43’43.9”S 72°28’51.4”W, 33masl, 15.xii.2013, coll. R.I. Madriz [1 ♀] GoogleMaps ; Río Negro Chaqueihua 0.5km N of intersection to Hornopirén N.P. 41°55'38.2"S 72°25'19.6"W, 39masl, 15.xii.2013, coll. R.I. Madriz & G.W. Courtney [4L, 1Ƌ, 1♀]; Río la Arena @ Puente la Arena 42°00’03.6”S 72°26’11.8”W, 32masl, 16.xii.2013, coll. R.I. Madriz [1P, 1♂] GoogleMaps ; Río Santa Juanita @ Puente Santa Juanita 42°07’38.1”S 72°27’54.8”W, 32masl, 16.xii.2013, coll. R.I. Madriz [2Ƌ] GoogleMaps ; Río Pichicolo southwest Hornopirén 41°59’07.7”S 72°33’51.7”W, 19masl, 17.xii.2013, coll. G.W. Courtney [3L]; Región XI (Aysén): Creek crossing @ highway 7 46°14’46.0”S 72°47’31.9”W, 273masl, 27.xii.2013, coll. R.I. Madriz [1Ƌ]; West of intersection to Caleta Tortel 47°46’48.1”S 73°19’00.2”W, 51masl, 26.xii.2013, coll. R.I. Madriz [11Ƌ, 3♀]; Creek @ road to Ventisquero Mont 6km south of intersection to Villa O’Higgins 48°02’58.7”S 73°07’52.7”W, 74masl, 25.xii.2013, coll. R.I. Madriz [1Ƌ]. GoogleMaps
Natural history. Habitat. All larvae were collected in water temperatures from 4 to 11°C, with most sites closer to 4°C. The riparian zone of inhabited streams varied from dense forest to sites with little to no riparian cover ( Fig. 3 View FIGURE 3 ). Collection sites typically had coarse gravel to rocky substrata, with submerged wood either sparse or abundant. Some sites had diagonal and mid-channel gravel bars. Substrata often contained abundant detritus. Stomach contents of captured specimens suggest Araucoderus larvae feed on fine particulate organic matter.
Captive reared larvae became increasingly active a few days prior to pupation, possibly in search of a suitable pupation site. During morning field collections, larvae were sometimes found crawling on the surface of the marginal substrate. Captive individuals became increasingly active during the morning from 0900–1100hrs. In the field, mature larvae were often found beside a rock that was much larger than the larva, sometimes partially buried in the contiguous sandy substrata ( Fig. 52 View FIGURES 46 – 53 ).
Resting behavior of adults. In both the field and laboratory, we observed no A. gloriosus adults resting on horizontal surfaces. The few specimens found on sloped surfaces used their tibial spurs as the main point of support. In the field, most adults were found on the underside of broad leaves in the riparian zone or among exposed roots of undercut river banks in the same general location as larvae ( Figs. 46–53 View FIGURES 46 – 53 ), sometimes more than 10 m away from the stream. Among the most common resting sites were the undersides of leaves of Gunnera sp., Chusquea sp., or Fuschia sp. A few individuals of both sexes were found under bridges, between the gabions armoring bridge abutments or on vertical surfaces of rocks and logs. A morphological study of the tarsal segments in A. gloriosus revealed similarities to those of Mischoderus Handlirsch, 1909 ( Friedemann et al. 2014) and suggests that specific tarsal attachment devices are not required for the observed resting behavior in Araucoderus and Mischoderus .
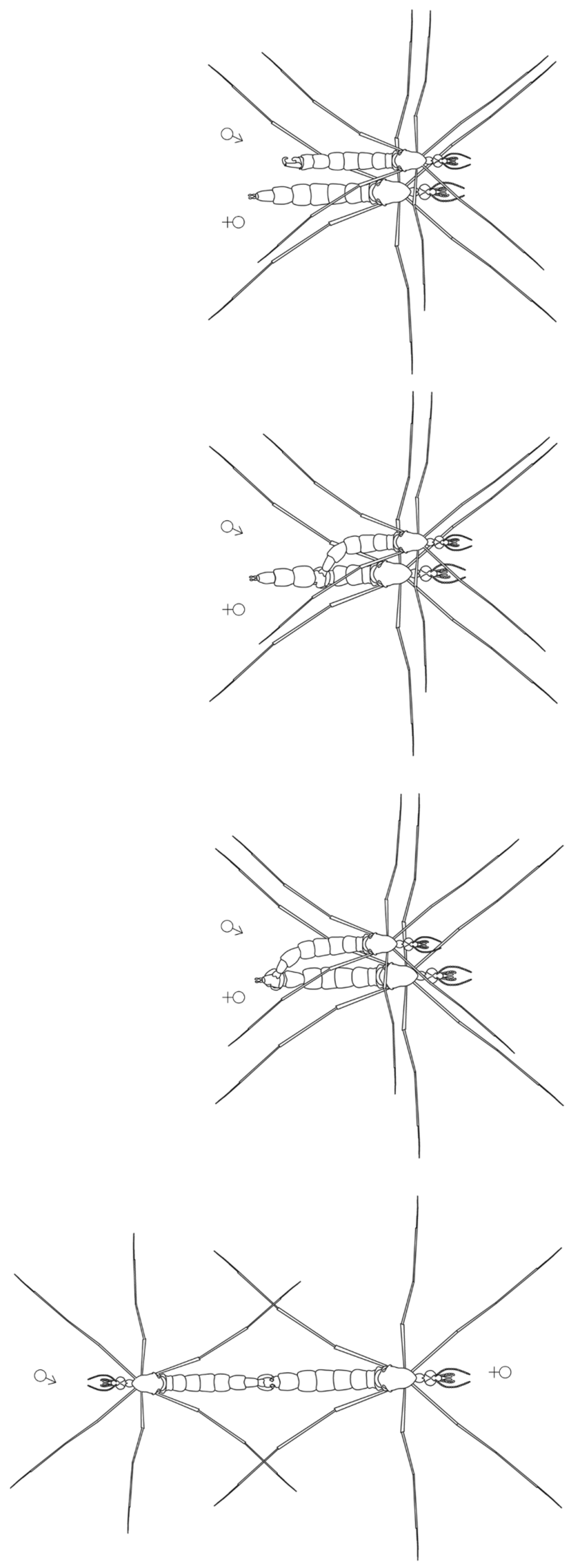
Mating behavior ( Fig. 54 View FIGURE 54 ). On December 13th 2013, a male and female of A. gloriosus were captured and kept alive in a clear container for further observation. Within 30 minutes of capture, the flying male collided several times with the vertically resting female, until coming to rest on the vertical wall of the enclosure side by side. Soon after, the male flexed the abdomen laterally, clasped the female abdomen at mid-length and guided its posterior end toward that of the female using an alternate constricting motion of the gonopods until their terminalia were aligned somewhat perpendicular. The male subsequently turned 180°, displaying a tail to tail copulation position. The copulation event was brief (10–20 seconds), followed by the male dislodging from the female. Neither individual showed further interest toward the other. The same female was observed copulating again several days after the first event. During the second copulatory event, three males were kept in the same enclosure.
During observed copulations, the apical tarsal segment of the foreleg was not used to hold the female, which disagrees with the general mechanism proposed by Hennig (1968). In fact, our observations suggest the female is restrained entirely by the gonopods. The only previous account of tanyderid mating position ( Borkent et al. 2008) suggested a generalized orientation in which the mating position after landing was tail to tail while both individuals are standing up. This agrees with our observations except that Araucoderus was unable to stand freely on a horizontal surface.
Our observations of mating in Araucoderus show some similarities with the copulatory behavior of Sycorax silacea Curtis, 1839 (Psychodidae) described by Jung ( Jung 1956 p.119) “… The male stands on the side of the female, with the body directed in the same direction as her. Suddenly he turns rapidly backwards with open claspers onto the female's abdomen. If the copulation succeeds, both animals stand for about 5 minutes calmly, forming a straight line with their bodies.” This observation may be a coincidence, but could be additional evidence supporting a close relationship between Tanyderidae and Psychodidae . Furthermore, Jung’s (1956) observations for S. silacea may also characterize the mating behavior of the Neotropical psychodid Aposycorax chilensis Tonnoir, 1929 , which exhibits similar behavior and morphology such as short flight pattern, resting in a hanging position and 90° rotation of the male genitalia ( Curler et al. 2015).
Oviposition. Clusters of eggs ( Fig. 4–6 View FIGURES 4 − 6 ) were laid on water droplets on the enclosure floor.
Night collections. Previous nocturnal collections of some tanyderids suggest a correlation of flight activity with sunset and evening hours ( Judd 2004). Furthermore, black- or incandescent lights can attract some species of Protanyderus Handlirsch (1909) , Protoplasa Osten Sacken, 1859 and Mischoderus ( Krzemiński & Judd 1997; Judd 2004; Courtney unpublished observations). During our 2013-2014 expedition, black-light sampling was conducted at different times throughout the night, in areas where immature and adult Araucoderus were present during the afternoon. However, these efforts yielded no adult specimens, suggesting that Araucoderus is not attracted to black lights. On one occasion, with no A. gloriosus adults approaching the light and no flight activity observed during sunset hours, a subsequent search of the marginal vegetation revealed nearly two dozen resting A. gloriosus adults. Based on our observations, Araucoderus adults may be most active during the morning hours.
Predation. Mid-air predation of adult Araucoderus was observed by a swallow ( Passeriformes : Hirundinidae ) and a dragonfly (Odonata: Anisoptera). Although no predation of larvae or pupae was observed, we found a dead mature pupa parasitized by over two dozen larval Phoridae .












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Araucoderus gloriosus ( Alexander, 1920 )
| Madriz, R. Isaí & Courtney, Gregory W. 2016 |
Radinoderus gloriosus
| Alexander 1929: 1 |
| Alexander 1927: 189 |
Tanyderus gloriosus
| Alexander 1920: 13 |