
Rhamma dawkinsi PrieTo & Lorenc-Brudecka, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4338.3.12 |
|
publication LSID |
lsid:zoobank.org:pub:AC0D026B-9A47-48AA-A632-B80243C1AC3F |
|
DOI |
https://doi.org/10.5281/zenodo.6041795 |
|
persistent identifier |
https://treatment.plazi.org/id/6D278785-D348-FFF1-FF60-F935900EFF17 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhamma dawkinsi PrieTo & Lorenc-Brudecka |
| status |
sp. nov. |
Rhamma dawkinsi PrieTo & Lorenc-Brudecka sp.nov.
( Figures 3, 4 View FIGURES 1 – 6 , 10, 11, 12 View FIGURES 7 – 12 )
Type material. Holotype male: COLOMBIA, Antioquia, Belmira, Páramo de Belmira , 3100 m, 05/01/2011, specimen number: RCCP m1309, CP Lep 0 0 99, GWOTI669-12 ; BOLD:ABX0547; C. Prieto Leg. The holotype is deposited in ICN-MHN.
Paratypes: 3 ♂ RCCP: COLOMBIA, Antioquia, Belmira, Páramo de Belmira , 3100 m, 05/01/2011, specimen numbers: m1303, m1304, m1307; C. Prieto leg. ; 2 ♂ RCCP: COLOMBIA, Antioquia, Belmira, Páramo de Belmira , 3100 m, 02/08/2015, specimen numbers: m1872, m1871; C. Prieto leg. ; 3 ♀ RCCP: COLOMBIA, Antioquia, Belmira, Páramo de Belmira , 3100 m, 05/01/2011, specimen numbers: m1305, m1306, m1308, C. Prieto leg.
This new species belongs to Rhamma shares the following combination of characters with all other species of Rhamma : 1) androconial scent brand appearing as an elongate streak bordering the upper vein of the discal cell apex on DFW ( Fig. 3 View FIGURES 1 – 6 ); 2) male genitalia with vinculum forming a 90 degree angle basally ( Fig. 10 View FIGURES 7 – 12 ); 3) vinculum strut strongly developed ( Fig. 10 View FIGURES 7 – 12 ); 4) ductus bursae short, robust, and with strongly developed lamella postvaginalis terminating in serrate, or multi-pronged configurations ( Fig. 12 View FIGURES 7 – 12 ); 5) fan shaped and dendritic signa ( Fig. 12 View FIGURES 7 – 12 ); 6) ductus bursae in dorsal or ventral aspect lightly sclerotized in the middle so that it appears to be transparent ( Fig. 12 View FIGURES 7 – 12 ); and 7) a ventral element associated with the 8th tergite in female genitalia (figure 3:C, J in Johnson 1992).
Diagnosis. Rhamma dawkinsi appears to be a sibling species of the sympatric R. adunca as both have a very similar ventral wing pattern. Rhamma dawkinsi differs from R. adunca ( Figs. 1, 2 View FIGURES 1 – 6 ) in having a smaller anal lobe and a longer and thinner androconial cluster. Additionally, R. dawkinsi is consistently smaller than R. adunca , and the upper margin of the hindwing turns down less abruptly than in R. adunca . Genital structures are practically indistinguishable ( Figs. 7, 8, 9 View FIGURES 7 – 12 ) as in the most of the species in the genus.
Description. Adult male wings. Mean forewing length 10.9 mm (measured from forewing apex to base at thorax); n = 3. Hindwing anal angle pointed, without tail at vein Cu2. Dorsal wing surface homogeneous silverblue, except for black margin extending from postmedial region of forewing and very narrow black margin (1–2 mm) on hindwing. No apparent orange scaling in anal angle. Ventral forewing surface gray with two thin, straight lighter lines ( Figs. 3, 4 View FIGURES 1 – 6 ). Conspicuous white band crossing medial area of ventral hindwing from costa to vein Cu2. Androconial cluster appearing as an elongate streak bordering ¼ of upper vein of discal cell apex on DFW.
Etymology. This species is named after the eminent English ethologist, evolutionary biologist, and popularizer of Science, Richard Dawkins, for his contributions to the public understanding of evolution and his fight against irrational thinking.
Biology. Males land on vegetation 1–2 m above the ground in the páramo-high Andean forest ecotone. Males appear to establish mating territories in the early afternoon around 1330 hours on the sunny edges of paths or on ridge tops. The spatial separation and the distance from the ground of the perching places of this species show a clear segregation from the individuals of R. adunca that fly on the same hilltops. Rhamma dawkinsi prefers small isolated bushes (1–2 m high) where males stay for a few minutes before leaving for another bush in a combination of perching and patrolling behaviour, whereas R. adunca males perch on leaves of small trees 4–5 m high. The immature stages, larval food plants, and adult nectar sources are unknown. Adults were captured in January and August.
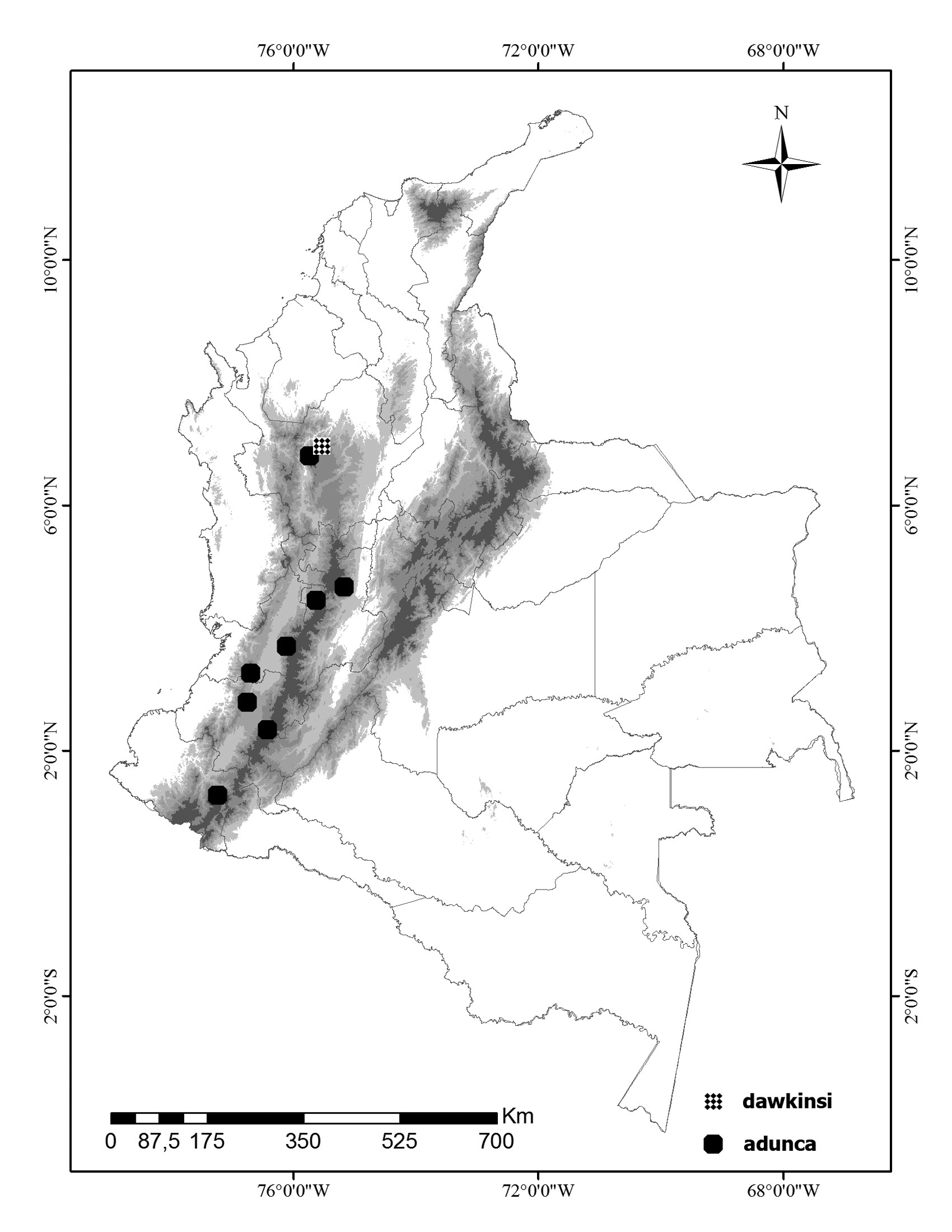
Distribution. The species is known only from the type locality, Páramo de Belmira in the central mountain range of the Colombian Andes, at 3100 m elevation ( Fig. 15 View FIGURE 15 ).
Remarks. Differences in size, wing pattern, and behaviour, as well as sympatry with the absence of intermediate individuals between both phenotypes and molecular comparisons, combine to show that R. adunca and R. dawkinsi , sp. n., are closely related, but different species. Although associating the sexes of many Eumaeini is difficult, Rhamma dawkinsi has a restricted geographical range and the wing pattern is almost identical in both sexes.
Molecular diagnostic characters. The intraspecific haplotype diversity in the available sequences (n = 3) was Hd = 3. Overall mean distance among the available sequences is 0.30%, maximum intraspecific distance is 0.31%. The lowest overall mean distance to another member of the genus is 2.98% to R. comstocki (eastern cordillera).
The molecular diagnostic characters when compared with the most similar species based on morphology are in the Figure 14. A View FIGURE 14 guanine in the position 453 of the mtDNA gen COI is diagnostic for this new taxon when compared with the 13 species recorded in Colombia by Prieto & Vargas (2016).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |