
Hexapanopeus klausruetzleri, Felder & Thoma, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5168.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:9D7A0DD5-630E-495E-BF61-55EEBB16D061 |
|
DOI |
https://doi.org/10.5281/zenodo.6916318 |
|
persistent identifier |
https://treatment.plazi.org/id/6C6A87D8-946D-FFBC-FF31-FB76FC9067D1 |
|
treatment provided by |
Plazi |
|
scientific name |
Hexapanopeus klausruetzleri |
| status |
sp. nov. |
Hexapanopeus klausruetzleri View in CoL n. sp.
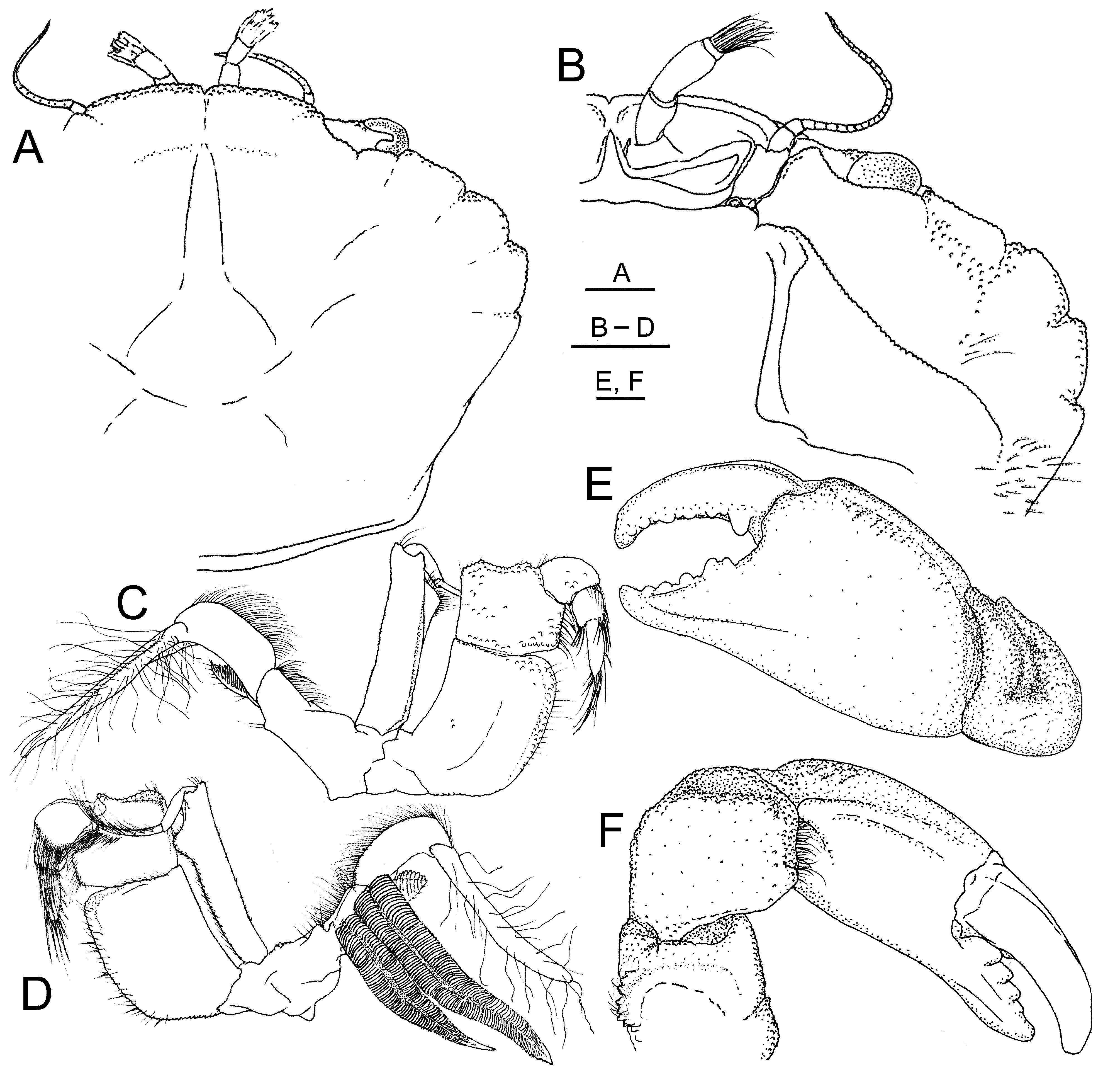
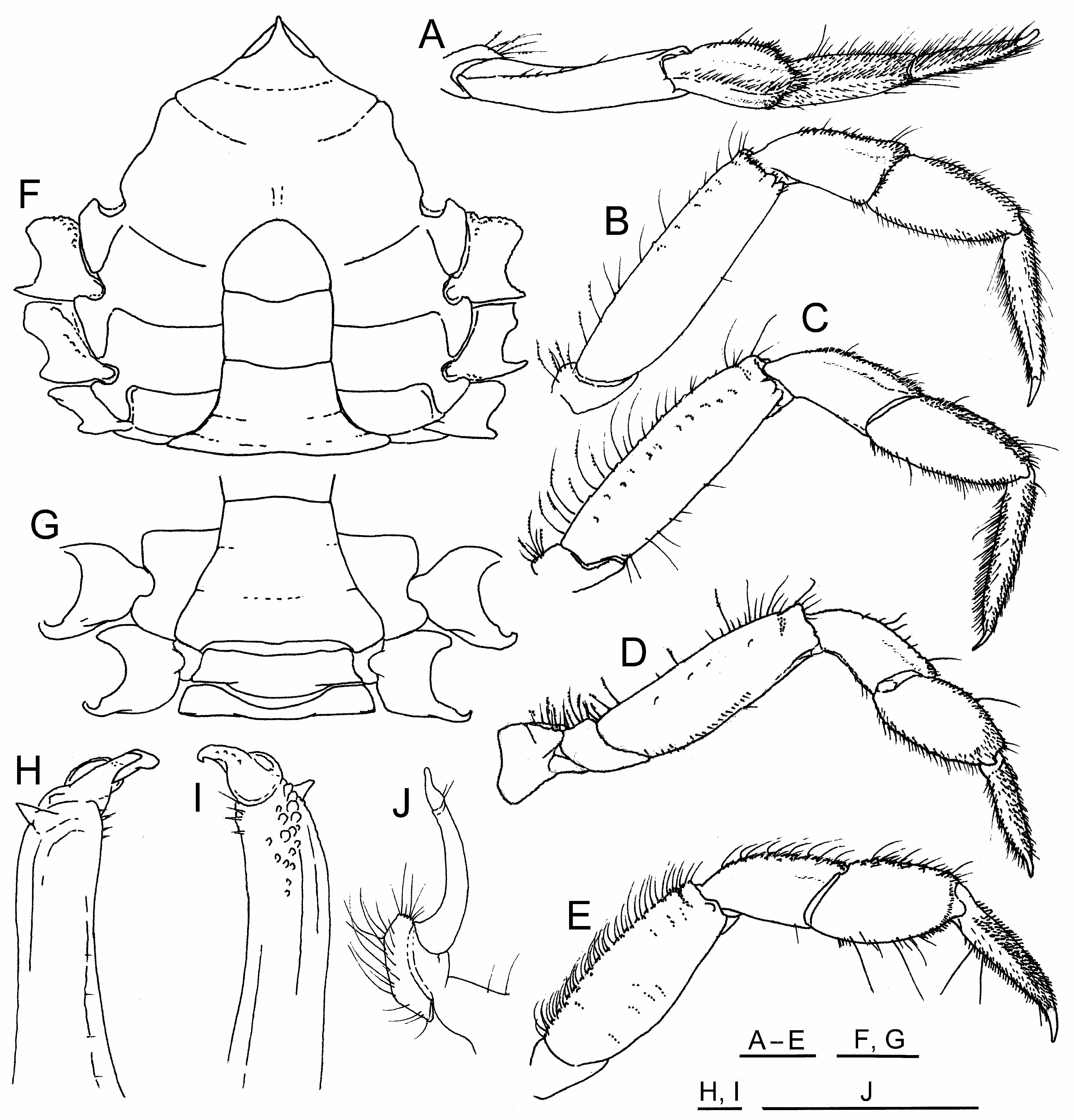
( Figs 1A–F View FIGURE 1 , 2A–J View FIGURE 2 , 3A–C View FIGURE 3 )
Hexapanopeus nov. sp. — Thoma et al. 2014: 89, table 1, 92, fig. 1.
Type material. Holotype: male, cw 9.9 mm, in dead conch shell on shallow subtidal sand flat behind reef break, off south end of South Water Cay , Belize, 16°48.682´N, 88°04.968´W, snorkeling, 1 m, 3 February 2011, coll. D. Felder, R. Lemaitre, J. Felder, S. Pecnik, & C. Tudge, USNM 1546469 About USNM (= ULLZ 12526 View Materials -A) GoogleMaps . Paratypes: 1 juvenile male, cw 7.2 mm, collection data same as for holotype, USNM 1661745 About USNM (= ULLZ 12526 View Materials -B) GoogleMaps ; 1 juvenile female, cw 5.2 mm, in eroded dead conch shell from deep patchy Thalassia and sponge bed, off northeast side of Twin Cays , Belize, 16°50.133´N, 88°05.852´W, snorkeling, 4 m, 21 April 2015, coll D. Felder, J. Scioli, K. Barkel, J. Felder, & C. Craig, USNM 1549828 About USNM (= ULLZ 16524 View Materials ) GoogleMaps .
Diagnosis. Carapace dorsal surface weakly convex, ovoid to weakly subhexagonal outline, wider than long (including anterolateral teeth), fronto-orbital width slightly exceeding 0.6 carapace width; front broadly convex with small median V-shaped notch separating shallow unimarginate lobes to each side, frontal margin of each lobe bearing slightly enlarged granules, lateral tooth of each frontal lobe obsolesent, lateral margin deflected to intersect antennal sinus; dorsal regions weakly defined, furrows of mesogastric, gastric, and anterior cardiac most evident; five anterolateral teeth, first tooth (outer orbital corner) subacutely angular, weakly separated from lobiform second by shallow depression of margin, third and fourth broadly lobiform to somewhat rounded or subrectangular, fifth small, subacutely angular. Third maxilliped merus distal margin sinuous, slightly produced to form small prominence near articulation with carpus. Cheliped merus superolateral margin with rough row of denticles, variably grouped to form serrate teeth, in major cheliped some forming sharp serrate teeth; carpus superior and lateral surfaces smooth to minutely granulate, microgranules forming weakly defined lines or surmounting weak elevations, supero-external surface marked by depression parallel to distal margin, internal margin with proximal angle weak, surmounted by line of low granules, distal angle stronger, forming blunt distally directed tooth. Major chela propodus mostly smooth to microgranulate, superior longitudinal crest weak, unarmed, adjacent external furrow shallow; dactylus opposable margin with slightly enlarged, lobiform basal tooth proximally. Ambulatory pereopods 2–5 relatively narrow, 2–4 subequal in size, merus length in all greater than 3 times greatest width, merus length approximating or slightly exceeding 1/2 carapace length; pereopod 5 smaller, propodus stouter than in other ambulatory pereopods, merus length not exceeding 1/2 length of carapace; merus in all ambulatory pereopods with superior margin unarmed, at most with dentiform granules variably concealed by plumose setae, inferior margin finely granulate; carpus superior margin with dense tract of short stout papilliform setae along crest; propodus superior margin with dense field of papilliform setae, dactylar-propodal locking mechanism not developed; dactylus stout proximally, inferior margin lacking subterminal prehensile tooth, superior margin densely covered by short papilliform setae, inferior margin with less dense slightly longer stiff simple setae. Male anterior thoracic sternum length (sternites 1–4) about 2/3 greatest width of fourth sternite (including episternites), eighth sternite exposed in subquadrate gap between lateral margin of flexed second pleonite and pereopod 5 coxa, exposure narrowly extending to anterior of condyle. Pleon of male with third through fifth pleonites fused, widest reach of first pleonite at its articulation with carapace, laterally rounded to articulation with second pleonite; telson subtriangularly rounded, widest in proximal one-third. Male first gonopod tip of highly modified panopeid form, subterminal tooth triangular, trunk in distal 1/4 of pleonal side bearing elongate field of heavy, rounded tubercles and granules extending to base of subterminal tooth. Male second gonopod less than one-third length of first gonopod. Applicable GenBank sequence accession numbers from Thoma et al. (2014) as follows for USNM 1546469 (= ULLZ 12526-A, holotype): (12S) KF683061 View Materials ; (16S) KF682952 View Materials ; (18S) KF682930 View Materials ; ( COI) KF682772 View Materials ; ( ENO) KF682722 View Materials ; (H3) KF682613 View Materials .
Description. Carapace ( Figs 1A, B View FIGURE 1 ; 3A–C View FIGURE 3 ) about 1.3 times wider than long, dorsal surface weakly convex, transverse edges of frontal lobes unimarginate, frontal width slightly exceeding 0.5 fronto-orbital width, frontoorbital width slightly exceeding 0.6 greatest carapace width, dorsal outline ovoid to slightly subhexagonal, dorsal regions overall weakly marked by furrows, marginal furrows of mesogastric, gastric, and anterior cardiac regions evident, most others obsolescent, dorsal surfaces of frontal regions less elevated than those to posterior, transverse ridges of granules weak to obsolescent, dorsal surfaces appearing mostly smooth with low cover of small granules, densest and largest granules developed near frontal and anterolateral margins; frontal margin overall broadly convex, small V-shaped median notch separating shallow unimarginate lobes to each side, each with transverse tract of slightly enlarged granules along frontal margin, lateral tooth obsolescent (weakly evident in small specimens), forming rounded corner, lateral margin of front deflected, angled to intersect antennal sinus just below mesial end of tooth on supraorbital margin; supraorbital margin granulate, forming low obtuse tooth above antennal sinus, median and lateral fissures forming distinct breaks in marginal granulation, margin convex between fissures. Anterolateral teeth well developed along distinctly convex arch of granulate margin, granules strong along anterior slopes of teeth; anterior two of five teeth broadly fused into subquadrate prominence, first (outer orbital corner) subacutely angular, very weakly separated by shallow depression of granulate margin from weakly lobiform second; third slightly produced at coarsely granulate anterior angle, lateral margin convexly rounded; fourth broadly triangular to subquadrate, angular anterior tip subacute, lateral margin weakly convex; fifth small but distinctly developed, subacutely angular. Posterolateral finely granulate with sparse cover of elongate plumose setae. Infraorbital margin ( Fig. 1B View FIGURE 1 ) granulate, forming strong subtriangular tooth mesially below base of eyestalk, margin sinuous laterally, ending in weakly produced infraorbital tooth. Pterygostomial and subhepatic regions with scattered low granules, coarsest near margins and along outer subhepatic region, lacking enlarged subhepatic tubercle, pterygostomial ridge distinctly granulate, slightly raised. Branchiostegite posterolaterally with sparse cover of elongate plumose setae, partially concealing ventrolateral margin above coxae of ambulatory legs.
Eyestalk stout, anteriorly with short precorneal patch of enlarged, raised granules. Antenna with long flagellum, peduncle with fused basal article elongate, rectangular, extending into orbital fossa.
Third maxilliped ( Fig. 1C, D View FIGURE 1 ) protopod elongate, distal surface weakly grooved to intersect ventral edge of carapace, bearing pair of unequal projections along margin internal to groove, proximal to small podobranch gill positioned adjacent to pair of much longer, lamellate, arthrobranch gills. Epipod thin, flattened, strap-like, bearing long simple setae along length. Endopod basis subtriangular; ischium broadly subrectangular, proximal end curved laterally to intersect basis, external surface mostly smooth with few small, granules densest marginally and in external tract along distomesial margin, weak evidence of longitudinal furrow medially, mesial margin with sparse fringe of simple setae; merus subquadrate, lateral margin near straight to very weakly concave, distolateral margin weakly angular to rounded, distal margin sinuous, produced to form distinctly raised tooth-like prominence near articulation with carpus, obliquely excavate distomesially to accommodate articulation and flexure of carpus, internal surface with short transverse tract of mesially directed setae extending into excavation below articulation of carpus; carpus short, stout, internal surface with distal fringe and distal submarginal field of elongate setae overlying propodus; propodus cylindrical, internal, superior, and inferior surfaces distally with submarginal and distal rows of elongate, stiff setae; dactylus elongate, digitiform, tapering distally, length about 1.5 times that of propodus, internal surface and inferior margin bearing short stiff setae, tip bearing dense tuft of long stiff setae approximating length of dactylus. Exopod weakly arched, elongate, narrowly subrectangular, internal edge of mesial margin produced to form strong rounded subtriangular projection in distal third, flagellum multi-articulate, bearing numerous long, distally directed setae.
Chelipeds (first pereopods) ( Figs 1E, F View FIGURE 1 ; 3A–C View FIGURE 3 ) somewhat unequal, texture mostly smooth or minutely granulate, few setae overall, slightly more setose along some margins of merus and carpus; ischium and merus with few plumose setae along tracts of enlarged granules on inner margins, small dense setal tuft near proximal inner margin of ischium; merus superolateral margin with roughly defined row of denticles, variably grouped to form serrate teeth, in major cheliped some forming enlarged, sharp serrate teeth, proximally with few plumose setae; carpus superior and lateral surfaces smooth to minutely granulate, microgranules forming weakly defined lines or surmounting elevations, supero-external surface marked by distinct depression parallel to distal margin, proximal to which weakly defined ridges of microgranules on supero-external surface, internal margin with proximal angle ill-defined, rounded, surmounted by line of low granules, distal angle stronger, produced to form blunt, subtriangular, distally directed tooth.
Major chela propodus mostly smooth to microgranulate, superior longitudinal crest of palm weak, unarmed, adjacent external furrow shallow; fixed finger of major chela subtriangular, exceeding ½ length of palm, inferior margin weakly convex, thick subacute tip curved upwards, opposable margin bearing five to six subtriangular to rounded teeth, proximal pair slightly fused, dark pigmentation extending proximally to base of finger but not onto palm; dactylus arched, slightly longer than fixed finger, opposable margin with slightly enlarged, lobiform, basal tooth proximally, distally with four to five somewhat rounded teeth separated by few smaller rounded denticles or granules, narrowing to subacute tip curved to cross to internal side of fixed-finger tip when flexed.
Minor cheliped propodus similar to that of major in form and length, palm less robust and slightly less elevated than that of major, dorsal microgranulation slightly coarser than that of major; fixed finger of minor chela closely resembling but slightly longer than that of major, opposable margin with six enlarged triangular to subtriangular teeth, distal three most closely grouped and least acute, dark pigmentation extending proximally beyond finger slightly onto distal extreme of palm; dactylus of minor chela resembling that of major, opposable margin lacking enlarged basal lobiform tooth, cutting edge dentition slightly weaker than in major chela, five to six teeth enlarged, proximal three subtriangular, distal lower and more rounded, subacute tip crossing to internal side of fixed-finger tip when flexed.
Ambulatory pereopods 2–5 relatively narrow, elongate, all of similar general form ( Fig. 2A–E View FIGURE 2 ; 3A–C View FIGURE 3 ); pereopods 2–4 subequal in size, merus length in each greater than three times greatest width, length of meri approximating or slightly exceeding 1/2 carapace length; pereopod 5 smaller, propodus relatively stouter than in other ambulatory pereopods, merus length not exceeding 1/2 that of carapace; ambulatory pereopod meri all with superior margin relatively unarmed, at most bearing a few small dentiform granules variably concealed by row of elongate, plumose setae, distal superior margin ending in low, blunt angle beyond subdistal notch, inferior margin finely granulate, at most with few setae, granules coarsest, densest proximally on pereopod 2; carpus superior margin with pubescence formed by dense tract of short stout papilliform setae along crest, few longer narrower setae, crest roughly paralleling weak to obsolescent secondary crest on superoposterior surface, secondary crest stronger on fourth pereopod than others; propodus superior margin with pubescence formed by dense field of papilliform seta, few longer narrower setae, inferior margin lined by primarily short, narrow, simple setae along with few longer; dactylar-propodal locking mechanism not developed; dactylus stout proximally, narrowing in distal half to weakly falciform corneous tip, inferior margin lacking subterminal, calcareous, prehensile tooth, superior margin densely covered by short papilliform setae forming broad tract of thick pubescence, inferior margin with less dense tract of slightly longer stiff simple setae, both margins with few additional long simple setae, dactylus terminating in weakly hooked, acute corneous tip.
Male anterior thoracic sternum length (sternites 1–4) longer than broad ( Fig. 2F, G View FIGURE 2 ), length from acute anterior apex to suture between fourth and fifth sternites about 2/3 greatest width of fourth sternite (including episternites), sternopleonal depression in fourth sternite not unusually sculpted or excavate to accommodate first gonopod tips below flexed pleon, median line originating from triangular depression at posterior of fourth sternite, narrowing to smooth very shallow groove bisecting fourth sternite, becoming obsolete to anterior; fifth sternite with small granuliform tubercle (part of press-button locking system) to each side of sternopleonal depression, each centered near 3/5 of distance from sixth sternite to fourth sternite; fourth and fifth episternites narrowly angular, subtruncate to anterior, subacutely rounded to posterior, sixth and seventh episternites slightly broader and more rounded posteriorly, eighth sternite exposed only in subquadrate gap between lateral margin of flexed second pleonite and pereopod 5 coxa, exposure narrowly extending to anterior of condyle on pereopod 5 coxa.
Pleon of male ( Fig. 2F, G View FIGURE 2 ) with third through fifth pleonites fused, first pleonite widest at articulation with carapace, rounded to articulation with second; second pleonite widest proximally near articulation with first, narrowing distally; fused third through fifth pleonites widest at lateral flange of third, weakly sinuous laterally, narrowing distally, width at articulation with sixth pleonite about half that at wide flange of third, vestiges of sutures between fused pleonites obscure; sixth pleonite subrectangular, slightly broadened distally before articulation with telson; telson subtriangularly rounded, widest in proximal third.
Male first gonopod tip ( Fig. 2H, I View FIGURE 2 ) of highly modified panopeid form, subterminal tooth triangular, terminal lobes not well-separated to produce trifid appearance typical of most panopeid crabs, trunk in distal 1/4 of pleonal side bearing elongate field of heavy, rounded tubercles and granules extending to base of triangular subterminal tooth, distolateral margin bearing several stiff setae proximal to terminal lobes. Penis emerging from pore on mesial surface of pereopod 5 coxa concealed beneath seventh episternite, extending beneath lateral flange formed by base of first gonopod. Male second gonopod less than one-third length of first gonopod, narrowing to subacute tip bearing several subterminal setae ( Fig. 2J View FIGURE 2 ).
Color. The carapace and pereopods are primarily golden brown to pale orange in presently limited materials, most dorsal surfaces being speckled with scattered dark reddish brown spots ( Fig. 3A–C View FIGURE 3 ). The pigment is overall darker on upper surfaces of chelipeds and ambulatory legs of larger specimens, especially the holotype, which is the largest. White borders the margins of most pereopod joints, and a distinct white spot is developed on the upper distal propodus on the cheliped, positioned immediately proximal to articulation of the movable finger. Fingers of the chelae vary in color from horn to reddish brown, with lighter tips. All pigmentation is pale in the most immature specimen.
Etymology. The species name is assigned in recognition of Klaus Ruetzler, world authority on marine sponge biology, who also led the establishment of the Smithsonian Marine Field Station at Carrie Bow Cay, Belize and for many years directed the Caribbean Coral Reef Ecosystems (CCRE) program based there. His encouragement and support of collaborators and colleagues brought many to focus years of work in Belize, including the senior author of this paper.
Size. Carapace widths ranged from 5.2 mm in the small female paratype to 9.9 mm in the male holotype.
Distribution. At present known only from tropical waters in the vicinity of South Water Cay and Twin Cays along the barrier reef of Belize.
Habitat. Thus far known from only high salinity shallow subtidal calcareous sand habitats (1–4 m depth), occupying dead gastropod shells on wash-over fans and back-reef flats of coral reefs, sometimes between patches of turtle grass and sponges.
Remarks. Systematic and taxonomic studies of western Atlantic panopeid crabs have been advanced over recent decades through detailed comparative studies of male gonopods ( Guinot 1967, 1968, 1969a –c, 1971, 1978, 1990; Martin & Abele 1986; Felder & Martin 2003). However, the advent of molecular genetic analyses has provided yet another powerful tool of particular value in understanding of phylogenetic relationships among otherwise very similar appearing panopeid crabs ( Schubart et al. 2000, Thoma et al. 2009; Thoma et al. 2014; Felder & Thoma 2020). Thoma et al. (2014: 98), on the basis of molecular genetic analyses, concluded that confirmed membership of Hexapanopeus s.s. was restricted to very few species, with the only two named species at that time being the western Atlantic H. angustifrons and H. paulensis Rathbun, 1930 . Their analysis also included several undescribed lineages, one of which is herein represented by H. klausruetzleri n. sp.
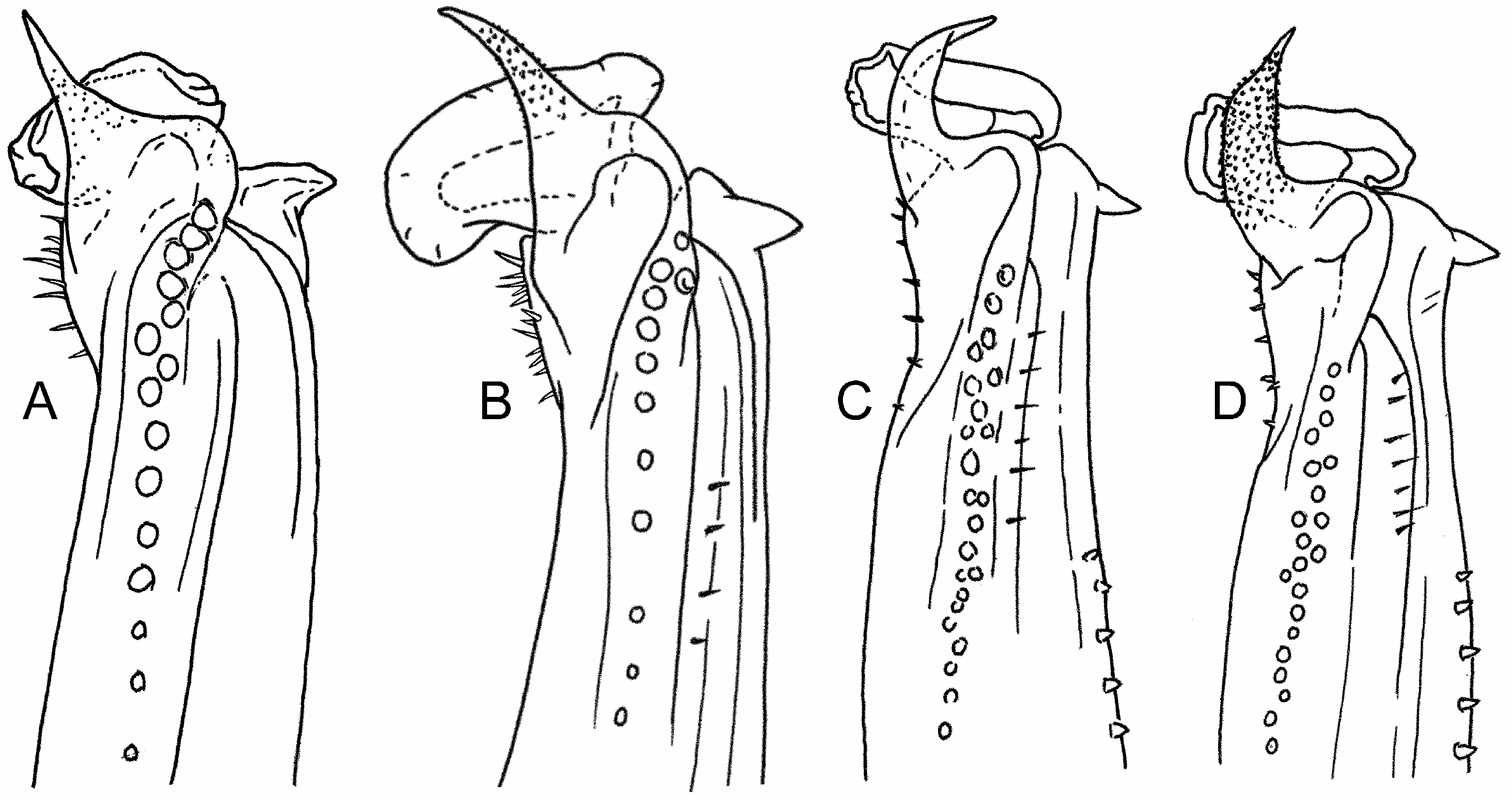
Morphologically, mature males of both the previously described congeners differ markedly in first gonopod morphology from that found in Hexapanopeus klausruetzeri n. sp. ( Fig. 2H, I View FIGURE 2 ), the first gonopods of H. paulensis and H. angustifrons having been previously illustrated by Williams (1965: figs 183D, E). Unfortunately, no mature female specimens of H. klausruetzleri n. sp. are presently available for description and comparative study of the fully developed female gonopore, a character that can also be of utility in panopeid crab separations. However, the previously described species of both sexes differ from H. klausruetzleri n. sp. in having a carapace front that is more truncate than arcuate, and in having the first and second anterolateral teeth distinctly separated ( Williams 1965: figs 170, 171) rather than weakly defined to either side of a shallow depression ( Fig. 1A, B View FIGURE 1 ). In addition, the anterolateral teeth are overall more angular and the carpus of the major cheliped is more roughly sculptured in both of the previously described species than in H. klausruetzleri n. sp. Sculpting of the major cheliped carpus in H. paulensis is especially distinct, with its dorsal surface typically bearing 8–15 well-defined knoblike tubercles. Sculpting of the carpus in H. angustifrons , while much less pronounced than in H. paulensis , is at least somewhat rougher than in H. klausruetzleri n. sp., the species to which it has a close sister relationship in molecular genetic analyses ( Thoma et al. 2014).
It appears that morphological separation of the closely related tropical Hexapanopeus klausruetzleri n. sp. and warm-temperate H. angustifrons can also be based upon additional characters, though presently limited material of the new species limits knowledge of their variability. Available specimens of H. klausruetzleri n. sp., exhibit stronger dentition of the cheliped merus, with its superolateral margin armed by row of denticles, some of which are grouped to form serrate teeth that are especially enlarged and sharply serrate on the major cheliped ( Fig. 1F View FIGURE 1 ). By contrast, this margin in H. angustifrons is armed by a low ridge ending in a blunt triangular tooth distally, bearing at most a low tubercle and one or two very weak subacute teeth along its length (TAMU/TCWC 2-2789). Additionally, H. klausruetzleri n. sp. is overall smoother in texture and ornamentation of the pereopods than is H. angustifrons . This is especially evident in the better-defined and more coarsely granulate carpal ridges in H. angustifrons , both on the chelipeds and the ambulatory pereopods (though not depicted in detail by Williams 1965: fig. 170).
The broadly angular, rounded, to subrectangular shape of the anterolateral carapace dentition in Hexapanopeus klausruetzleri n. sp. is somewhat similar to that seen in subadult to adult stages of Eurypanopeus abbreviatus (Stimpson, 1860) , E. ater Rathbun, 1930 , and Panopeus americanus Saussure, 1857 . In both E. abbreviatus and E. ater , the carapace anterolateral teeth are more consistently evident as shallow lobes, with the third being distinctly subrectangular and most of its lateral margin straight, while the anterior and posterior ends form rounded corners. By contrast, the anterolateral teeth in H. klausruetzleri n. sp. are in general more produced and lobiform, the third with an anterior corner that is distinctly more angular than the rounded curve or corner of its posterior slope. In P. americanus , the third and fourth anterolateral teeth, while similar in breadth to those of H. klausruetzleri n. sp., are by contrast more strongly produced anteriorly, the posterolateral margins being arched conspicuously to create broad lobes, sweeping the tip of tooth more toward the anterior than in H. klausruetzleri n. sp. However, mature males of these species are also readily distinguished from those of H. klausruetzleri n. sp. by the morphology of their first gonopods ( Fig. 4A–D View FIGURE 4 ).
In addition to the herein described new taxon, we have identified a number of other panopeid and closely related pseudorhombilid crab species from the vicinity of South Water Cay, Carrie Bow Cay, Twin Cays, and Dangriga, Belize. While three of these were previously among materials utilized in molecular analyses by Thoma et al. (2014: table 1), the others represent species not reported previously from Belize (Table I). At least some of those listed likely share habitats with Hexapanopeus klausruetzleri n. sp., although there is little present understanding of the latter’s habitat limitations. At least two of the species listed among these, Acantholobulus schmitti (as noted in the Introduction) and Scopolius nuttingii ( Rathbun, 1898) , represent materials that warrant additional study, with the present materials from Belize perhaps representing in whole or part undescribed taxa.
Our sampling efforts for panopeid and closely related pseudorhombilid crabs from the coast of Belize have been far from exhaustive, geographically or in terms of represented habitats, and all of our present collections are from shallow to intertidal waters. Previous reports from nearby shallow waters of Quintana Roo (Markham et al. 1990; Mejía-Ortíz et al. 2008), elsewhere in adjacent Caribbean habitats ( Hernández-Ávila et al. 2007; Poupin 2018), suggest that both Eurypanopeus abbreviatus and Eurypanopeus dissimilis ( Benedict & Rathbun, 1891) might also be expected to range into coastal waters of Belize. The shallow burrowing species Eucratopsis crassimanus (Dana, 1852) , known to populate northwestern Caribbean habitats similar to those sampled in Belize (MartinezIglesias and Raso (1999), should also be expected in these waters.

| R |
Departamento de Geologia, Universidad de Chile |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Xanthoidea |
|
Family |
|
|
Genus |
Hexapanopeus klausruetzleri
| Felder, Darryl L. & Thoma, Brent P. 2022 |
Hexapanopeus
| Thoma, B. P. & Guinot, D. & Felder, D. L. 2014: 89 |