
Austrocheirus isasii
|
publication ID |
https://doi.org/ 10.5281/zenodo.204710 |
|
DOI |
https://doi.org/10.5281/zenodo.5620784 |
|
persistent identifier |
https://treatment.plazi.org/id/6C127405-F63A-1478-FF39-FDADF8C7FF41 |
|
treatment provided by |
Plazi |
|
scientific name |
Austrocheirus isasii |
| status |
|
Austrocheirus isasii gen. et sp. nov.
Figures 2 View FIGURE 2 A, D, H, J, 3–7, 9C, F
Etymology. The generic name is derived from the Greek words austros (south) and cheirus (hand, manus); in allusion to the non-atrophied manus of this taxon which constitutes the first example of a medium-sized Cretaceous abelisauroid with this condition. The specific name is in honor of Mr. Marcelo Isasi, for his outstanding work in the last 25 years as a technician in palaeontology and discoverer of the holotype specimen of Austrocheirus .
Holotype. MPM-PV 10003, specimen comprising several incomplete mid-caudal vertebrae, left metacarpal III, proximal left manual phalanx of digit III, distal end of left tibia, fragmentary left metatarsal III, and some fragments of pedal phalanges.
Diagnosis. Medium-sized theropod (7–8 meters in total length; Table 2 View TABLE 2 ) diagnosed by the following combination of characters (autapomorphies indicated by an asterisk) ( Fig. 2 View FIGURE 2 ): (1) metacarpal III with a dorsoventrally tapering shaft towards its proximal end* and (2) posteriorly displaced collateral tendon fossae located at the same level of the proximal end of distal condyles*; (3) tibia with a slightly convex posterior surface at the distal end; (4) metatarsal III with a very well-developed ventral scar for the insertion of the M. gastrocnemius ; (5) pedal phalanges with a conspicuous longitudinal crest delimitating the dorsal margin of the distal collateral tendon fossae*.
Locality and horizon. Hoyada Arroyo Seco locality (49°57’17’’ S, 72°05’31’’ W), close to the western coast of the Leona River and four kilometers south of the De los Hornos Hill, southwest Santa Cruz Province, Patagonia, Argentina ( Fig. 1 View FIGURE 1 ). Pari Aike Formation, Late Cretaceous (early Maastrichtian: Kraemer & Riccardi 1997; Novas et al. 2004).
Note on the association of the holotype specimen. The bones of MPM-PV 10003 were found closely associated within a two square metres area and match one another perfectly in size. MPM-PV 10003 was found in a green to grey mudstone. No other tetrapod remains were found mixed with it, and the next closest specimen was a fragmentary sauropod found around 300 metres away. Additionally, as discussed below, most of the remains, especially the metacarpal and distal tibia, possess ceratosaurian apomorphies. Thus, all the bones of the holotype of Austrocheirus must correspond to a single individual.
Description. With regard to the controversial issue of theropod manual digit homology, it must be noted that metacarpal III of Austrocheirus will be compared with metacarpal III of tetanurans (= metacarpal IV sensu Wagner & Gauthier 1999; see Xu et al. 2009), because their development seems to be determinate and controlled by the same set of morphogenetic genes ( Burke & Feduccia 1997; Wagner & Gauthier 1999).
The preserved transverse width of the distal end of the tibia of Austrocheirus is of almost the same size as that of Ekrixinatosaurus (7–8 metres in total length: Calvo et al. 2004) and Sinraptor (7.2 metres in total length: Currie & Zhao 1993) (see Table 2 View TABLE 2 ). Thus, Austrocheirus constitutes a medium-sized animal of 7–8 meters in total length, and is thus larger than the abelisaurids Aucasaurus and Majungasaurus ( Coria et al. 2002; Burch & Carrano 2008) ( Table 2 View TABLE 2 ), both of which possess atrophied hands.
Metacarpal III. A left metacarpal III without its proximal end and a damaged distal articular trochlea is preserved in the holotype of Austrocheirus ( Fig. 3 View FIGURE 3 A–G). This element is identified here as metacarpal III because of the presence of an elongated but robust shaft and collateraly projected shelves directly above the collateral tendon pits. By contrast, metacarpals I and II of other theropods are proportionally more robust elements, with the proximal transverse expansion closer to the distal trochlea, whereas metacarpal IV is always a more reduced bone in other basal theropods. In addition, the morphology of the available metacarpal of Austrocheirus closely resembles that of metacarpal III of Ceratosaurus (USNM 4735).
Metacarpal III of Austrocheirus is relatively elongated, being proportionally more gracile than that of derived abelisaurids (e.g. Aucasaurus , Carnotaurus: Coria et al. 2002 ; MACN-Pv-CH 894, Bonaparte et al. 1990), but resembling basal saurischians (e.g. Herrerasaurus , Dilophosaurus : PVSJ 373, 380, Sereno 1993; UCMP 37302, Welles 1984) and non-abelisaurid ceratosaurians (e.g. Ceratosaurus, Limusaurus ; USNM 4735, Gilmore 1920; Xu et al. 2009). Nevertheless, metacarpal III of Austrocheirus is more robust than that of basal tetanurans (e.g. Szechuanosaurus, Allosaurus , Acrocanthosaurus: Gao 1993 ; Madsen 1976; Currie & Carpenter 2000). In addition, the elongated metacarpal III of Austrocheirus also resembles the proportions of metacarpal II of the basal ceratosaur Berberosaurus ( Allain et al. 2007).
The proximal end of the bone is sub-rectangular in cross-section ( Fig. 3 View FIGURE 3 F). The preserved portion of the shaft indicates that the area for contact with metacarpal II was restricted to the proximal end of the bone ( Fig. 3 View FIGURE 3 A). The shaft is dorsoventrally compressed, being two times lower than the height of the distal articular condyles ( Fig. 3 View FIGURE 3 A, B). In other theropods a dorsoventral compression of the shaft towards its proximal end is absent (e.g. Allosaurus , Dilophosaurus, Liliensternus, Carnotaurus, Aucasaurus , Torvosaurus , Megaraptor , Acrocanthosaurus: Madsen 1976 ; UCMP 37302; MB R2175; MACN-Pv-CH 894; Coria et al. 2002; Galton & Jensen 1979; MUCPv 341; Currie & Carpenter 2000), but the metacarpal shaft of Austrocheirus tapers dorsoventrally towards the proximal end of the bone. In Carnotaurus (MACN-Pv-CH 894) and Aucasaurus ( Coria et al. 2002) the shaft of metacarpal III is as tall as the distal end, with a strongly dorsoventrally expanded proximal end. Despite the bizarre morphology of the hand of these South American abelisaurids, this condition resembles that of other theropods. By contrast, in Austrocheirus the shaft is lower than the distal end of the bone and tapers proximally. Accordingly, we consider this morphology to be autapomorphic for Austrocheirus ( Fig. 2 View FIGURE 2 A–C). The base of a conspicuous proximolateral process is present, resembling other theropods (e.g. Dilophosaurus, Ceratosaurus : UCMP 37302; USNM 4735) in which this process bears the articular surface for metacarpal IV ( Fig. 3 View FIGURE 3 C–E).
The dorsal margin of the distal articular condyles protrudes directly above the collateral ligament pits ( Fig. 3 View FIGURE 3 C, D, G), a condition which also seems to be present in the metacarpus of some ceratosaurians such as Ceratosaurus and Berberosaurus ( Gilmore 1920; Allain et al. 2007). The dorsal flexor fossa is laterally and medially flanked by each of these ridges. The dorsal flexor fossa of Austrocheirus is poorly defined and shallowly excavated ( Fig. 3 View FIGURE 3 C–D), as also occurs in the metacarpals of other ceratosaurs such as Ceratosaurus (USNM 4735, Gilmore 1920) and Berberosaurus ( Allain et al. 2007). By contrast, well-excavated dorsal flexor fossae, proximally bounded by a high transverse lip, are present in “coelophysoids” (e.g. Coelophysis , Dilophosaurus : QG 1; UCMP 37302) and basal tetanurans (e.g. Allosaurus , Megaraptor: Madsen 1976 ; MUCPv 341) or are completely absent in derived coelurosaurians ( Rauhut 2003). The ventral surface of the shaft exhibits a proximolaterally directed median ridge, located proximal to the distal end of the bone. The distal end of metacarpal III of Austrocheirus is pulley-shaped (i.e. the distal condyles are separated by a wide intercondylar groove) ( Fig. 3 View FIGURE 3 G), contrasting with the rounded distal end present in Limusaurus ( Xu et al. 2009) and abelisaurids (e.g. Aucasaurus , Carnotaurus: Bonaparte et al. 1990 ; Coria et al. 2002). The distal end of the metacarpal is strongly asymmetric, with a lateral condyle more proximodistally extended than the medial condyle, as in most theropods. The transition between the medial distal condyle and the shaft is gradual, whereas in basal tetanurans (e.g. Torvosaurus, Allosaurus , Acrocanthosaurus: Galton & Jensen 1979 ; Madsen 1976; Currie & Carpenter 2000) this transition forms a sharp angle. The distal collateral tendon pits are shallow and poorly defined, as occurs in derived abelisaurids (e.g. Aucasaurus , Carnotaurus: Coria et al. 2002 ; Bonaparte et al. 1990), but contrasting with “coelophysoids” (e.g. Dilophosaurus , Coelophysis : UCMP 37302; QG 1) and tetanurans (e.g. Torvosaurus , Allosaurus: Galton & Jensen 1979 ; Madsen 1976) in which they are well excavated and delimited. In contrast to other theropods (e.g. Allosaurus , Dilophosaurus, Liliensternus , Torvosaurus , Megaraptor , Acrocanthosaurus: Madsen 1976 ; UCMP 37302; MB R2175; MACN-Pv-CH 894; Coria et al. 2002; Galton & Jensen 1979; MUCPv 341; Currie & Carpenter 2000), the collateral tendon pits of Austrocheirus are proximally located, being situated at the proximal-most level of the distal articular condyles ( Fig. 2 View FIGURE 2 A-C, 3A-B). The collateral tendon pits are asymmetrically arranged, with the medial pit more ventrally situated than the lateral one.
Manual phalanges. A fragmentary manual phalanx is preserved, being a proximodistally short element as occurs in digit III of other theropods, thus probably belonging to one of the most proximal elements of left digit III ( Fig. 3 View FIGURE 3 H–L). This phalanx is stout and bears poorly defined and shallow collateral tendon pits, as also occurs in abelisaurids with reduced forelimbs such as Aucasaurus and Carnotaurus ( Coria et al. 2002; Bonaparte et al. 1990). However, in Austrocheirus these pits are strongly proximodistally extended, occupying almost half the entire length of the phalanx ( Fig. 3 View FIGURE 3 H–I). The distal articular condyles are asymmetric, with the lateral condyle more distally extended than the medial one ( Fig. 3 View FIGURE 3 L). The distal median intercondylar groove is poorly excavated ( Fig. 3 View FIGURE 3 K), in contrast with the pulley-like distal end present in “coelophysoids” (e.g. Dilophosaurus , Coelophysis : UCMP 37302; QG 1) and tetanurans (e.g. Allosaurus , Deinonychus: Madsen 1976 ; Ostrom 1969).
Tibia. The distal end of the left tibia is present, and preserves a damaged medial distal process but lacks a portion of the lateral distal process ( Fig. 4 View FIGURE 4 ). The tibia is not fused to the astragalus, contrasting with adult specimens of other basal neotheropods (e.g. Xenotarsosaurus, Masiakasaurus , Coelophysis: Rowe & Gauthier 1990 ; Tykoski 2005). The cross-section of the shaft is oval, with a transverse main axis ( Fig. 4 View FIGURE 4 F). The anterior surface of the shaft, directly above the reception area for the ascending process of the astragalus, exhibits an oblique, low, conspicuous, and rounded ridge. The posterior surface of the tibia of Austrocheirus is convex, a feature widely distributed among basal theropods and also present in Berberosaurus ( Allain et al. 2007) and Ceratosaurus (Madsen & Welles 2000) ( Fig. 2 View FIGURE 2 D–E). By contrast, a concave posterior surface of the distal end of the tibia is exhibited by Velocisaurus (MUCPv-41), Elaphrosaurus (MB unnumbered), Ekrixinatosaurus (MUCPv-294), Xenotarsosaurus (MACN-Pv-CH 1468), and Masiakasaurus ( Carrano et al. 2002) ( Fig. 2 View FIGURE 2 F–G). Furthermore, in some abelisaurids, including Ekrixinatosaurus (MUCPv-294) and Xenotarsosaurus (MACN-Pv-CH 1468), the depressed posterior surface of the distal tibia is bounded medially and laterally by two conspicuous vertical ridges, a feature absent in Austrocheirus , Velocisaurus (MUCPv-41), Elaphrosaurus (MB unnumbered), and Berberosaurus ( Allain et al. 2007).
The area for the reception of the ascending process of the astragalus is represented by a depressed and sub-rectangular scar on the anterior surface of the distal end of the bone. As occurrs in other basal abelisauroids (e.g. Velocisaurus ; MUCPv-41), this subrectangular scar is medially delimited by a vertical tuberosity ( Fig. 4 View FIGURE 4 A, B), which we consider homologous of the facet for the ascending process of the astragalus of non-abelisauroid dinosaurs. By contrast, in non-abelisauroid theropods (e.g. Zupaysaurus, Coelophysis , Dilophosaurus, Ceratosaurus , Piatnitzkysaurus , Torvosaurus , Neovenator , Sinraptor , Allosaurus: Ezcurra & Novas 2007 ; Raath 1990; UCMP 77270; Welles & Long 1974; PVL 4073; Britt 1991; Brusatte et al. 2008; Currie & Zhao 1993; Madsen 1976) the facet for the reception of the ascending process is represented by a conspicuous anteriorly raised step oblique to the longitudinal axis of the bone. In particular, the surface for reception of the ascending process of Austrocheirus is sub-rectangular in anterior view and well defined, as also occurs in most abelisauroids, such as Ozraptor ( Long & Molnar 1998; sensu Rauhut 2005), Elaphrosaurus (MB unnumbered), Xenotarsosaurus ( Martinez et al. 1986) , Velocisaurus (MUCPv-41) Masiakasaurus ( Carrano et al. 2002) , Skorpiovenator ( Canale et al. 2009) , Aucasaurus ( Coria et al. 2002) , and unnamed remains from the Late Jurassic of Africa ( Rauhut 2005). This surface extends along most of the width of the distal end of the bone, a condition also present in basal abelisauroids and coelurosaurians. The proximal half of the surface is almost flat and gently slopes posteriorly.
A sub-triangular median vertical ridge is present on the distal half of this articular surface, a condition also documented in Elaphrosaurus (MB unnumbered), Velocisaurus (MUCPv-41), Ozraptor , and unnamed abelisauroids from the Late Jurassic of Africa ( Rauhut 2005). Interestingly, this ridge seems to imbed within a vertical groove on the posterior surface of the ascending process of the astragalus ( Agnolin et al. 2005; i.e. an unnamed abelisauroid astragalus from the Early Cretaceous of Australia), suggesting that some kind of tibiotarsal interlocking was present. In the basal tetanuran Megalosaurus, Benson (in press) described a vertical ridge that divides the astragalar scar for the reception of the ascending process of the astragalus, probably articulating with a notch or groove on the posterior surface of the ascending process of the astragalus as occurs in Torvosaurus and Allosaurus ( Britt 1991; Madsen 1976). However, the vertical crest of basal tetanurans differs from that of abelisauroids in being an extremely thin, dorsoventrally reduced, and medially displaced structure. Furthermore, the pit for the reception of this ridge in Allosaurus and Torvosaurus is oblique and positioned at the base of the ascending process of the astragalus, contrasting with the groove of abelisauroids which is longitudinally extended on the entire posterior surface of the ascending process.
Metatarsal II?. Only a fragment of the mid-shaft of this element is preserved ( Fig. 5 View FIGURE 5 E, F). It is interpreted as a probable fragment of the shaft of the metatarsal II because it is thinner than the available metatarsal III and does not exhibit lateral curvature as occurs in metatarsal IV of dinosauriforms and, in particular, abelisauroids ( Novas 1996; Carrano et al. 2002; Carrano 2007). The metatarsal is oval in cross-section, with an almost flat anterior surface and a strongly convex posterior surface. The probable metatarsal II of Austrocheirus is wide and robust, which seems to differ from the extremely narrow metatarsal II of noasaurids (e.g. Masiakasaurus , Noasaurus: Carrano et al. 2002 ). Otherwise, the condition of Austrocheirus resembles that of other basal theropods (e.g. Dilophosaurus, Ceratosaurus, Allosaurus, Majungasaurus : UCMP 37302; Madsen & Welles 2000; Madsen 1976; Carrano 2007).
Metatarsal III. The distal third of a left metatarsal III, lacking the medial distal condyle, is preserved ( Fig. 5 View FIGURE 5 A–D, G, H). The shaft is slightly laterally bowed. The proximalmost preserved portion of the shaft is trapezoidal in cross-section, with the medial margin dorsoventrally taller than the lateral one. The medial surface of the bone is almost flat along most of its length; however, close to the distal end of the bone the medial surface becomes slightly convex. The medial surface is proximodorsally delimited by a sharp flange. This flange becomes distally a sharp edge which reaches the distal end of the metatarsal. The flat medial surface represents the extensive articulation with metatarsal II ( Fig. 5 View FIGURE 5 B). Conversely, the lateral surface of the shaft is convex along the entire preserved length of the bone. Close to the proximalmost preserved portion of the shaft, a poorly developed lateral tuber is present ( Fig. 5 View FIGURE 5 A, B, D). The convex lateral surface suggests that the contact between metatarsals III and IV was not as proximodistally extensive as that between metatarsals II and III, as is the case in Majungasaurus ( Carrano 2007) . In some abelisaurids, including Aucasaurus and Majungasaurus , two ventral ridges extend on the ventral surface of the shaft and meet each other distally. This V-shaped structure is not observed in Austrocheirus , but instead a large and well-developed scar is present, probably representing the insertion area of the Mm. gastrocnemii ( Carrano & Hutchinson 2002) ( Fig. 2 View FIGURE 2 H). This ventral tuber is located at mid-width of the transverse axis of the shaft. The tuber and the surface directly proximal to it are slightly ornamented by small furrows and pits. At the distal end of the bone, the dorsal extensor fossa is absent, a condition contrasting with that observed in other theropods (e.g. Dilophosaurus, Liliensternus, Allosaurus : UCMP 37302; MB R2175; Madsen 1976). The distal condyles of the metatarsal are dorsoventrally low, with its dorsal and ventral margins continuous with the metatarsal shaft in lateral view, a condition also present in other ceratosaurians (e.g. Ceratosaurus, Elaphrosaurus, Masiakasaurus , Majungasaurus: Gilmore 1920 ; Janensch 1925; Carrano et al. 2002; Carrano 2007). The distal condyles are ventrally separated by a wide and shallow intercondylar groove ( Fig. 5 View FIGURE 5 H). A circular, large, and well-defined collateral tendon fossa is preserved on the lateral distal condyle ( Fig. 5 View FIGURE 5 A).
Pedal phalanges. Only incomplete distal ends of non-ungual pedal phalanges are present ( Fig. 5 View FIGURE 5 I–M). These fragments resemble pedal phalanges rather than manual elements in having distal collateral tendon pits not dorsally displaced, anteroposteriorly wide, and sub-oval. Moreover, the distal articular surface of the phalanges is transversely wider than dorsoventrally deep, an opposite condition to that of manual elements. The distal condyles are separated from each other by a shallow intercondylar groove. The distal collateral tendon pits are wide, deep, oval, and poorly delimited anteriorly, posteriorly, and ventrally. The distal flexor fossae are poorly excavated and faintly defined. Two incomplete distal ends of pedal phalanges present a conspicuous crest that delimitates the dorsal margin of the distal collateral tendon fossae ( Fig. 5 View FIGURE 5 I–K), a feature not observed in other known theropod that we are aware (e.g. Ilokelesia , Velocisaurus, Majungasaurus, Aucasaurus, Elaphrosaurus : MCF-PVPH 36; MUCPv-41; Carrano 2007; MCF-PVPH 236; MB R2175) ( Fig. 2 View FIGURE 2 J–M).
Caudal vertebrae. Fragmentary remains of vertebral centra and incomplete neural arches of distal or mid-caudal vertebrae are preserved ( Fig. 6 View FIGURE 6 ). The articular surfaces of the caudal vertebrae are broader than tall and amphyplatan ( Fig. 6 View FIGURE 6 J, K). The centra do not bear pneumatic features. The available remains of neural arches ( Fig. 6 View FIGURE 6 A–I) exhibit elongate and posterolaterally oriented postzygapophyses and possess welldeveloped and thick neural spines. The neural spines are strongly dorsally projected above the dorsal margin of the postzygapophyses. In constrast to Majungasaurus ( O’Connor 2007) , the available caudal vertebrae of Austrocheirus show short and less caudally extended postzygapophyses. The preserved caudals of Austrocheirus lack hyposphene-hypantrum articulations, as occurs in other theropods in which these structures are restricted to the anterior caudal vertebrae. The postzygapophyses are close to each other, being separated by a transversely thin median notch. The neural canal seems to have been narrow, as suggested by the preserved dorsal walls. In contrast to the mid-caudal vertebrae of Majungasaurus ( O’Connor 2007) , the epipophyses are completely absent in Austrocheirus .
Internal bone morphology and ontogenetic stage. The state of preservation of metacarpal III allows us to observe features of its internal morphology in the region where it is broken ( Fig. 7 View FIGURE 7 ). The trabeculae of spongy bone are widely extended, restricting the cortical bone to almost 30% of the transverse diameter of the shaft. The cortical bone possesses a high density of secondary osteons on its inner half. By contrast, the outer half of the cortical bone exhibits a low density of secondary osteons, which is not consistent with a rapid growth stage. Radial vessels are not common in the cortical bone, contrasting with a juvenile stage ( Hayashi et al. 2009). A distinct and poorly vascularized lamellar-zone bone is present in the outermost cortex, indicating the external fundamental system (EFS) ( Cormack 1987) and suggesting a slow rate of bone accretion ( Chinsamy-Turan 2005). At least four sets of lines of arrested growth (LAGs) are observed in the most peripheral region of the cortical and lamellar bone. The presence of an EFS indicates that the holotype of Austrocheirus was an adult individual, but the absence of complete remodeling of cortical bone by secondary osteons suggests that it was not an old animal ( Sander 2000; Erickson 2005; Xu et al. 2007; cf. histological ontogenetic stage 12 of Klein & Sander 2008).
The fusion of the astragalus to the distal end of the tibia has been described by previous authors ( Rowe & Gauthier 1990; Tykoski & Rowe 2004; Tykoski 2005) as a feature present in adult specimens of non-tetanuran neotheropds. In particular, Tykoski (2005) found that the fusion between the astragalus and tibia is one of the later events that took part in the ontogeny of “coelophysoids” (ontogenetic stage 9 of Tykoski [2005]). In the holotype of Austrocheirus , the proximal tarsals are not fused to the tibia. The latter is in agreement with the evidence provided by the internal bone structure, and would suggest that this specimen represents a young adult individual at the time of death (sensu Hayashi et al. 2009), or alternatively that this kind of fusion did not take part during the ontogeny of Austrocheirus .
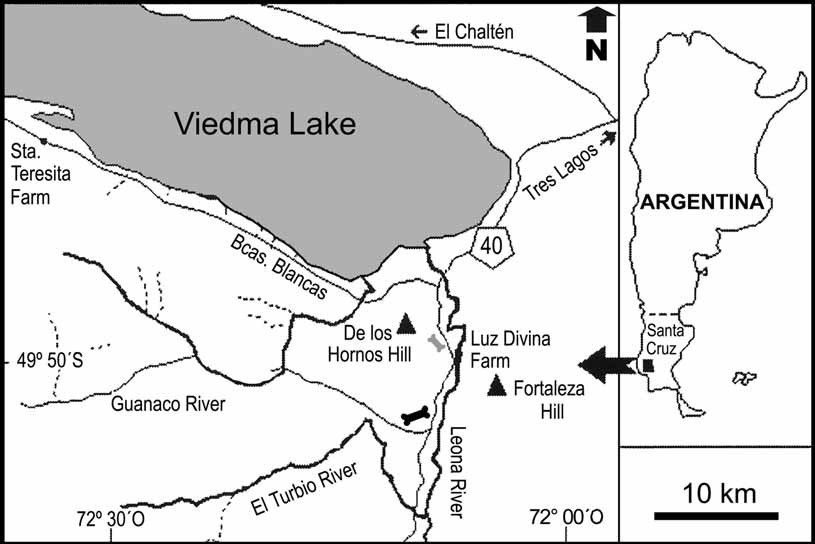
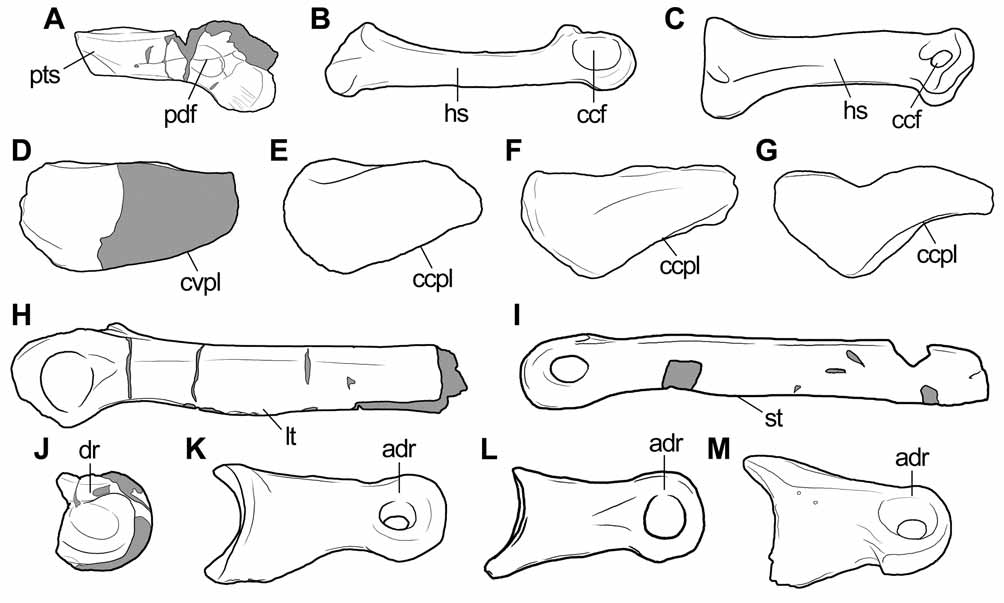
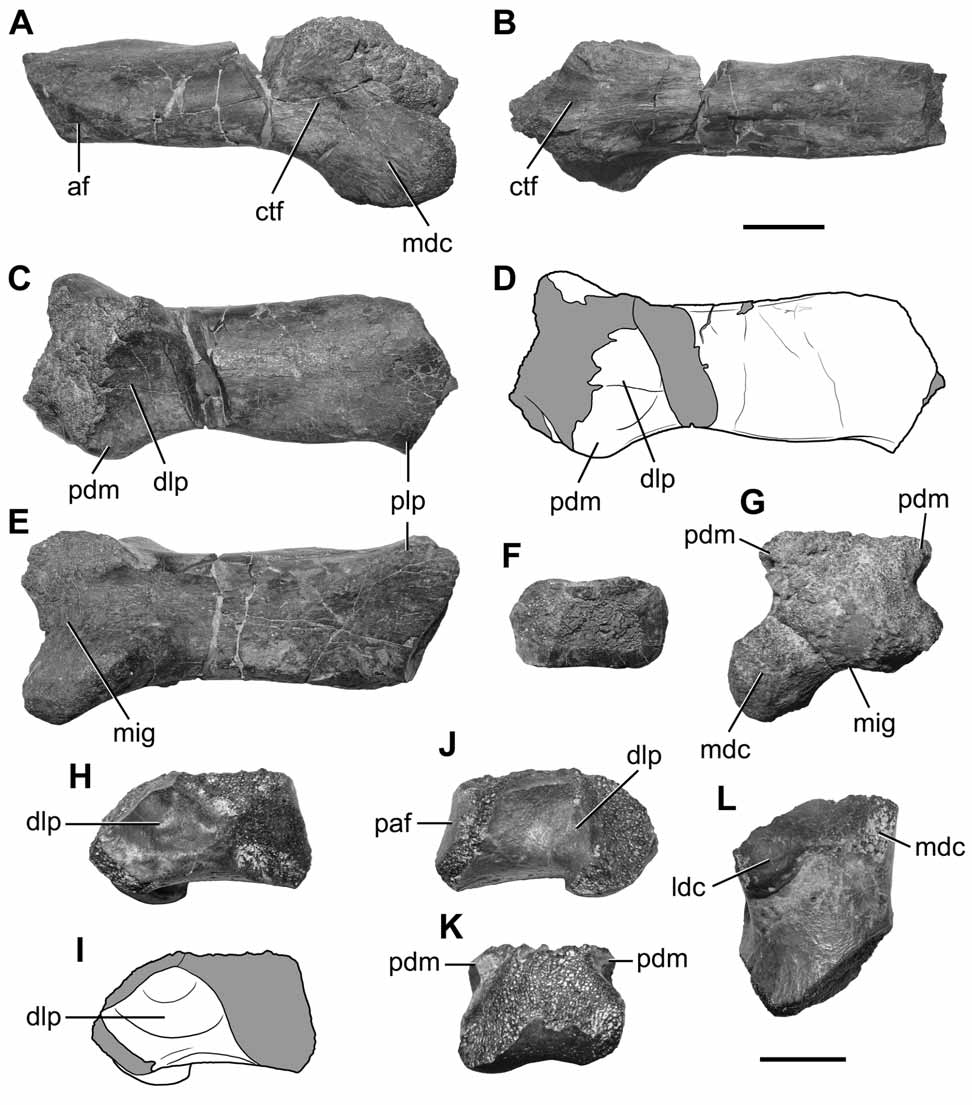
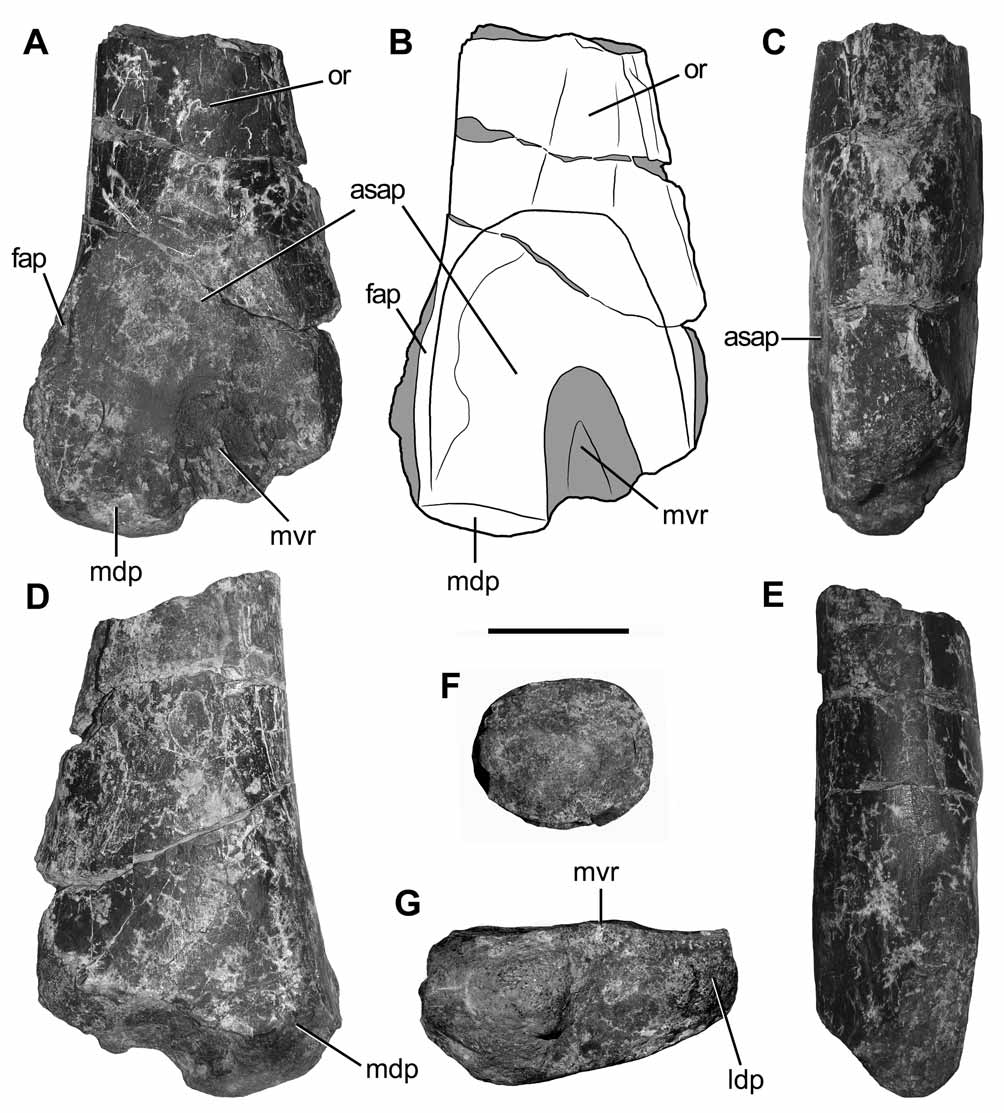


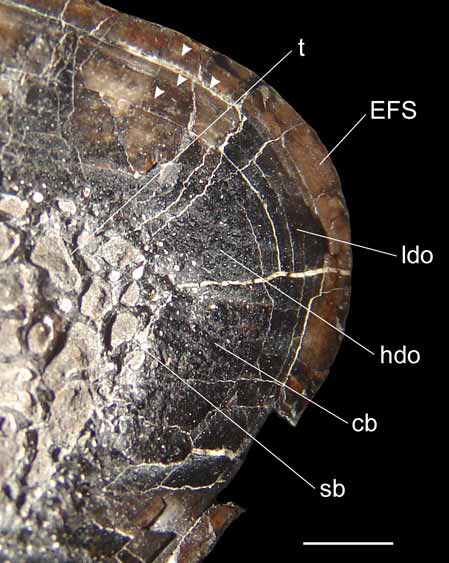
TABLE 2. Comparative measurements of metacarpal III and distal tibia of Austrocheirus isasii gen. et sp. nov. and several neotheropod taxa. Abbreviations: LMtcIII, length of metacarpal III; TWDT, transverse distal width of tibia. Tibiae ratio depicts the proportion between the TWDT of Austrocheirus (100 %) and that of each taxa.
| Taxa | TWDT | LMtcIII | Tibae Ratio | Source |
|---|---|---|---|---|
| Austrocheirus | > 183 | >107.73 | -- | This paper |
| Dilophosaurus | 80.3 | 80.8 | <43.8 % | UCMP 37302 |
| Ceratosaurus | 140 | 66.0 | <76.5 % | Gilmore 1920 |
| Berberosaurus | 82 | -- | <44.8 % | Allain et al. 2007 |
| Elaphrosaurus | 104 | -- | <56.8 % | MB unnumbered |
| Velocisaurus | 18 | -- | <9.8 % | MUCPv-41 |
| Masiakasaurus | 28 | -- | <15.3 % | Carrano et al. 2002 |
| Xenotarsosaurus | 134 | -- | <73.2 % | MACN-Pv-CH 1468 |
| Ekrixinatosaurus | 183 | -- | <100 % | MUCPv-294 |
| Majungasaurus | 70.2 | -- | <38.4 % | Carrano 2007 |
| Carnotaurus | -- | 29.0 | -- | MACN-Pv-CH 894 |
| Piatnitzkysaurus | 130 | -- | <71.0 % | PVL 4073 |
| Sinraptor | 195 | 151.8 | <106.5 % | Currie & Zhao 1993 |
| Mapusaurus | 288 | -- | <157.4 % | Coria & Currie 2006 |
| Tyrannosaurus | 312 | 104 | <170.5 % | Brochu 2003 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.