
Crenidorsum aroidephagus Martin & Aguiar
|
publication ID |
https://doi.org/ 10.5281/zenodo.4620270 |
|
persistent identifier |
https://treatment.plazi.org/id/687E87C9-D32A-FFA1-D270-FCACC503DD9F |
|
treatment provided by |
Plazi |
|
scientific name |
Crenidorsum aroidephagus Martin & Aguiar |
| status |
sp. nov. |
Crenidorsum aroidephagus Martin & Aguiar View in CoL sp. nov. ( Figs 1-3 View FIGURE 1 View FIGURES 2 3 )
Crenidorsum View in CoL sp. L. Nakahara, 1981a: 336; Paulson & Kumashiro, 1985: 110, 119.
PUPARIUM ( Fig. 1 View FIGURE 1 ). Habitus. Scattered under leaves, not in distinct feeding groups. Puparia do not secrete any visible dorsal or periferal wax coating, being cryptic when feeding and silvery when the adults have emerged. Shortly before emergence, the eyes of the adult can be clearly seen through the puparial cuticle, along with the developing body structure ( Paulson & Kumashiro 1985: 119, Fig. 3 View FIGURES 2 3 F).
Margin. Outline elongateoval, 0.801.10 mm long, 0.500.80 mm wide, 1.40 1.65 times longer than wide, generally widest at abdominal segment I/II (n=25). Anterior and posterior marginal setae present, fine, similar to each other, up to length of operculum. Margin evenly crenate, typically 1416 rounded teeth occupying 0.1 mm of margin (n=43); marginal teeth without glandular bases; margin not modified at thoracic and abdominal tracheal openings.
Dorsum. Chaetotaxy comprises setae that vary in length markedly, even within samples (see Table 1 View TABLE 1 ), ranging from short and stout, with sharply acute apices, to very long and hairlike: single pairs of cephalic (10300 m), mesothoracic (typically 530 m), metathoracic (typically 530 m), 8th abdominal (most 50 m) and caudal (5100 m) setae usually present (n=44), but see comments. A pair of distinct longitudinal folds leads from lateral to cephalic setae to abdominal segment V and, together with a marginconcentric fold anterior to cephalic setae, define the submedian area of the puparium, often as a distinct shallow rhachis; longitudinal folds slightly and variably scalloped, especially thoracically ( Fig. 2 View FIGURES 2 3 ). Longitudinal moulting suture slightly crimped in appearance ( Fig. 2 View FIGURES 2 3 ), reaching puparial margin, dividing the cephalic marginconcentric fold into two sections; transverse moulting sutures almost reach longitudinal subdorsal folds. Dorsal disc generally finely corrugate with very closely paired geminate pore / porettes distributed in an even pattern over dorsal disc; in addition to 24 geminate pore / porettes on each of abdominal segments IIVII, between the longitudinal folds, clusters of 24 simple thickrimmed pores are also present just mesal to the folds, with similar pores, sometimes clustered, also unevenly distributed adjacent to the folds cephalothoracically ( Fig. 2 View FIGURES 2 3 ). Meso/ metathoracic and abdominal segmentation distinct submedially, abdominal segment VII only slightly reduced in length medially, pockets distinct. A shallow shaped elevation is evident surrounding the vasiform orifice in many individuals (particularly clear in Fig. 1 View FIGURE 1 , and see Fig. 3 View FIGURES 2 3 ), defining a caudal furrow which terminates at the caudal setal bases, not the puparial margin. Vasiform orifice subcircular, almost fully occupied by the roundedtrapezoidal operculum. Lingula usually folded with its head between posterior edge of operculum and rim of vasiform orifice, appearing crescentshaped ( Fig. 3 View FIGURES 2 3 ), with its apical setae directed upwards; when unfolded ( Fig. 1 View FIGURE 1 ) the lingular head is seen to be globose, similar in size to operculum, finely setose and with acute apical setae around 30 m long.
Ven te r. Cuticle diaphanous and smooth; thoracic tracheal folds completely unmarked, except as very slight differential staining in some postemergence pupal cases; caudal tracheal folds slightly marked by very fine stippling, posterior to vasiform orifice (between posterior abdominal spiracles). Ventral abdominal setae almost underlying eighth abdominal setae, very fine and typically 25 30 m. Middle and hind legs each with a tiny basal seta usually just discernible, no longer than its own base. Each leg with an apical disc, usually termed “adhesion pad”; antennal bases placed anteromesal to fore legs.
MATERIAL EXAMINED. HOLOTYPE puparium, Madeira, Funchal, on Epipremnum aureus , 16 March 1999 (Aguiar) ( BMNH). PARATYPES: 28 puparia, Madeira, same data as holotype ( BMNH, ICLAM, USNM); 42 puparia, São Martinho, on Anthurium hybrid, February & November 1997; November 1998 (Aguiar) ( BMNH, ICLAM); 8 puparia, Madalena do Mar, on Philodendron sp., 26 October 1998 (Aguiar) ( ICLAM); 22 puparia, Germany, Berlin Botanic Gardens, on Philodendron gloriosum , March 2001 (Baufeld) ( BBA, BMNH, USNM); 1 puparium, Belize, Cayo, Chiquibul Forest Reserve, 23 km Las Cuevas Monkey Tail Branch, on undetermined aroid vine, 0 5 March 1996 (Martin) ( BMNH); 1 puparium, Cayo, Chiquibul Forest Reserve, Las Cuevas, on Araceae , 0 3 March 1996 (Martin) ( BMNH); 9 puparia, 1 thirdinstar larva, Brasil, S.P.,?Campinas, on Syngonium sp., received March 1983 (via Lourencão) ( BMNH); 8 puparia, Costa Rica, San Jose, on? Philodendron sp., 22 January 1983 (Martin) ( BMNH); 5 puparia, Hawaii, Honolulu, Foster Botanical Gardens, on Philodendron sp., 10 April 2001 (Kumashiro & Dooley) ( APHIS, BMNH). OTHER MATERIAL: 4 puparia, 1 thirdinstar larva, Costa Rica, Heredia Prov., 15 km north of Puerto Viejo, on? Philodendron sp., 0 3 February 1983 (Martin) ( BMNH); 1 parasitised puparium, Heredia Prov., 10 km north of Puerto Viejo, on Matisia ochrocalyx (Bombacaceae) , 0 2 February 1983 (Martin) ( BMNH); 12 puparia, 1 thirdinstar larva, 1 secondinstar larva, Panama, Canal Zone, Barro Colorado Island, on? Philodendron sp., 13 March 1983 (Martin) ( BMNH).
DISTRIBUTION. Neotropical Region Belize, Brasil, Costa Rica, Panama; Pacific Region American Samoa, Hawaii; Palaearctic Region Germany (under glass), Madeira; Nearctic Region California, Florida.
RECORDED HOST PLANTS. Araceae : Anthurium crenatum , A. lindenianum , A. longissimum , A. spp.; Culcasia scandens ; Epipremnum aureus , Monstera deliciosa var. borsigiana ; Nephthytis sp.; Philodendron gloriosum , P. spp.; Rhaphidophora korthalsii ; Spathiphyllum sp.; Syngonium podophyllum , S. cf. salvadorense, S. spp.; Xanthosoma sagittifolium . [ Bombacaceae : Matisia ochrocalyx ].
ETYMOLOGY. The specific name reflects the wide range of aroid hosts colonised by this whitefly.
COMMENTS. The puparium of this species displays little phenotypic plasticity, except in the dorsal setae. These vary greatly in length, and precisely which setae become much enlarged (in any individual) appears random, although usually enlargement occurs in pairs (see Table 1 View TABLE 1 ). In other specimens, some setal pairs are extremely short, again usually in pairs. Within the invasive populations, away from the Neotropics, the mesothoracic setae are always visibly present, but those from some Neotropical samples have the mesothoracic pair almost, or entirely, undetectable. Although it is considered that samples devoid of visible mesothoracic setae are most likely to be conspecific with those which clearly do possess them, the slight doubt leads to the decision to omit from the paratype series those samples without evident mesothoracic setae. A single parasitised specimen, damaged but apparently C. aroidephagus , collected from Matisia ochrocalyx (Bombacaceae) in Costa Rica is present in BMNH. In Madeira, several puparia collected from Philodendron sp. and Epipremnum aureus were parasitised by Cales noacki (Howard) (Eulophidae) and others, collected from an Anthurium hybrid, by Encarsia hispida (De Santis) (Aphelinidae) .
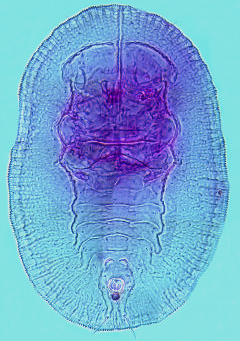
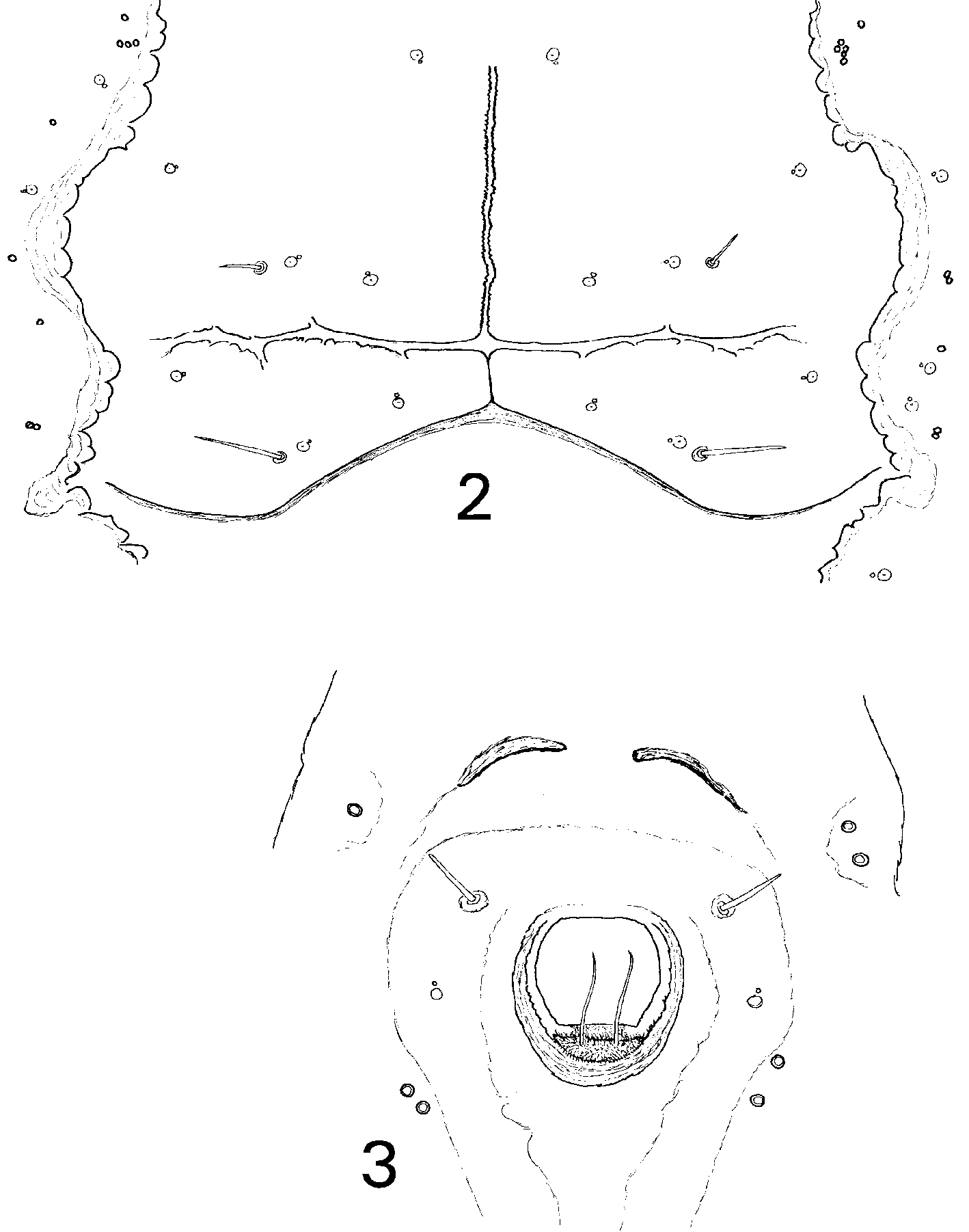
TABLE 1. Puparial seta measurements of Crenidorsum aroidephagus Martin & Aguiar sp. nov. from Madeira (µm).
| Sp.no | Cephalic | Mesothor. | Metathor. | 8th abd. | Caudal | Ventral abd. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | | | | 57.5 | 5 | 27.5 | |||||||
| 2 | | 13.8 | 17.5 | 5 | 12.517.5 | 3052.5 | |||||||
| 3 | | 511.3 | 11.3 | 12.5 | 56.3 | 25 | |||||||
| 4 | 35 | 12.5 | | 7.5 | 52.567.5 | 2527.5 | |||||||
| 5 | 283.1295.8 | 109.7249.9 | 272.9290.7 | 90125 | 112.5 | 27.5 | |||||||
| 6 | 13.8 | | | 7.5 | 13.817.5 | 27.5 | |||||||
| 7 | | | | | | 2526.3 | |||||||
| 8 | | 12.5 | | 1516.3 | 2027.5 | 2526.3 | |||||||
| 9 | 12.5 | 12.5 | 17.5 | 1017.5 | 11.3 | 27.5 | |||||||
| 10 | 28.8 | | | 16.3 | 15 | 25 | |||||||
| 11 | | | | | | 30 | |||||||
| 12 | | | | | 78.8 | | |||||||
| 13 | 260.1 | 21.3 | 36.3 | 21.327.5 | 58.8102.5 | 25 | |||||||
| 14 | | | 8.817.5 | 57.5 | 7.5 | 22.5 | |||||||
| 15 | 23.8 | | 15 | 10 | 20 | 25 | |||||||
| 16 | | | | | 105130 | 25 | |||||||
| 17 | | 10 | 1522.5 | 17.521.3 | 5058.8 | 23.8 | |||||||
| 18 | 11.3 | 5 | 5 | 6.3 | 7.5 | 25 | |||||||
| 19 | 2027.5 | 12.515 | 2527.5 | 22.5 | | 25 | |||||||
| 20 | | 12.5 | 6.310 | 12.513.8 | 1012.5 | 26.3 | |||||||
| 21 | 8.8 | 7.5 | 8.8 | 10 | 7.5 | 28.8 | |||||||
| 22 | 15 | 12.515 | 13.8 | 10 | 15 | 25 | |||||||
| 23 | 30 | 15 | 37.5 | 20 | | 28.8 | |||||||
| 24 | | | | | 22.5 | | |||||||
| 25 | 17.5 | 1015 | 12.5 | 6.37.5 | 15 | 1022.5 | |||||||
| 26 | 28.826.3 | 13.8 | 17.5 | 12.516.3 | 22.5 | 25 | |||||||
| 27 | | | | | 12.5 | 27.5 | |||||||
| 28 | 20 | 12.5 | 15 | 15 | | 27.5 | |||||||
| 29 | 12.5 | 6.3 | 11.3 | 5 | 8.8 | 30 | |||||||
| 30 | 55 | 15 | 35 | | 103.8117.5 | 21.3 | |||||||
| 31 | | | 15 | 10 | 3.8 | 27.5 | |||||||
| 32 | 13.816.3 | 7.5 | 17.5 | 10 | | | |||||||
| 33 | 34 | | 6.3 | | 11.3 | | 15 | 11.3 | 10 | 10 | 5 | 25 | |
| 34 | 6.3 | 11.3 | 15 | 10 | 5 | | |||||||
| 35 | | 6.3 | | 5 | 10 | 28. 8 | |||||||
| 36 | | 10 | 13.817.5 | 11.3 | 27.538.8 | | |||||||
| 37 | 311.1321.3 | 20.4 | 191.3280.5 | 25.5 | 53.668.9 | | |||||||
| 38 | 280.5 | 10 | 115225 | 42.5 | | 28.8 | |||||||
| 39 | 336357 | 5082.5 | 293.3300.9 | 8087.5 | 185 | 27.5 | |||||||
| 40 | 1021.3 | 6.3 | 15 | 6.3 | 41.3 | | |||||||
| 41 | 316.2 | 18.8 | 270.3285.6 | 16.335 | 75120 | 2532.5 | |||||||
| 42 | 323.9351.9 | 22.527.5 | 152.5153.8 | 62.563.8 | 152.5156.3 | | |||||||
| 43 | 265.2275.4 | 2530 | 200 | 52.5 | 98.8102.5 | | |||||||
| 44 | 316.2321.3 | 23.825 | 60 | 2531.3 | 97.5105 | 35 |
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Crenidorsum aroidephagus Martin & Aguiar
| Martin, J. H., Aguiar, A. M. F. & Baufeld, P. 2001 |
Crenidorsum
| Paulson 1985: 110 |
| Nakahara 1981: 336 |