
Jogoya, Anker, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5263.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:35D6B99A-B03B-460C-A450-1F3E929D8A94 |
|
DOI |
https://doi.org/10.5281/zenodo.7805352 |
|
persistent identifier |
https://treatment.plazi.org/id/655987C0-FFBA-FFFD-FF44-F99211DAF993 |
|
treatment provided by |
Plazi |
|
scientific name |
Jogoya |
| status |
gen. nov. |
Jogoya gen. nov.
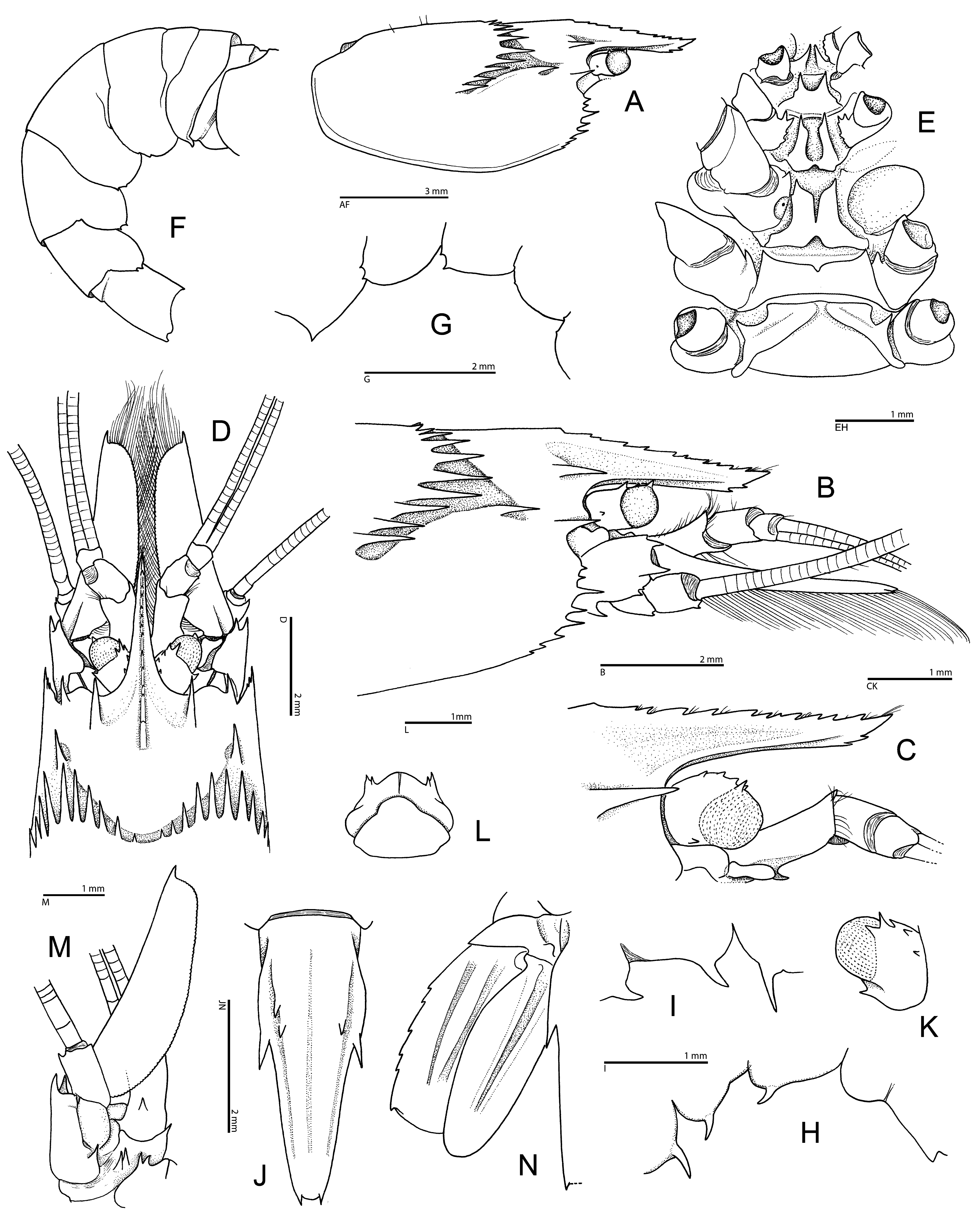
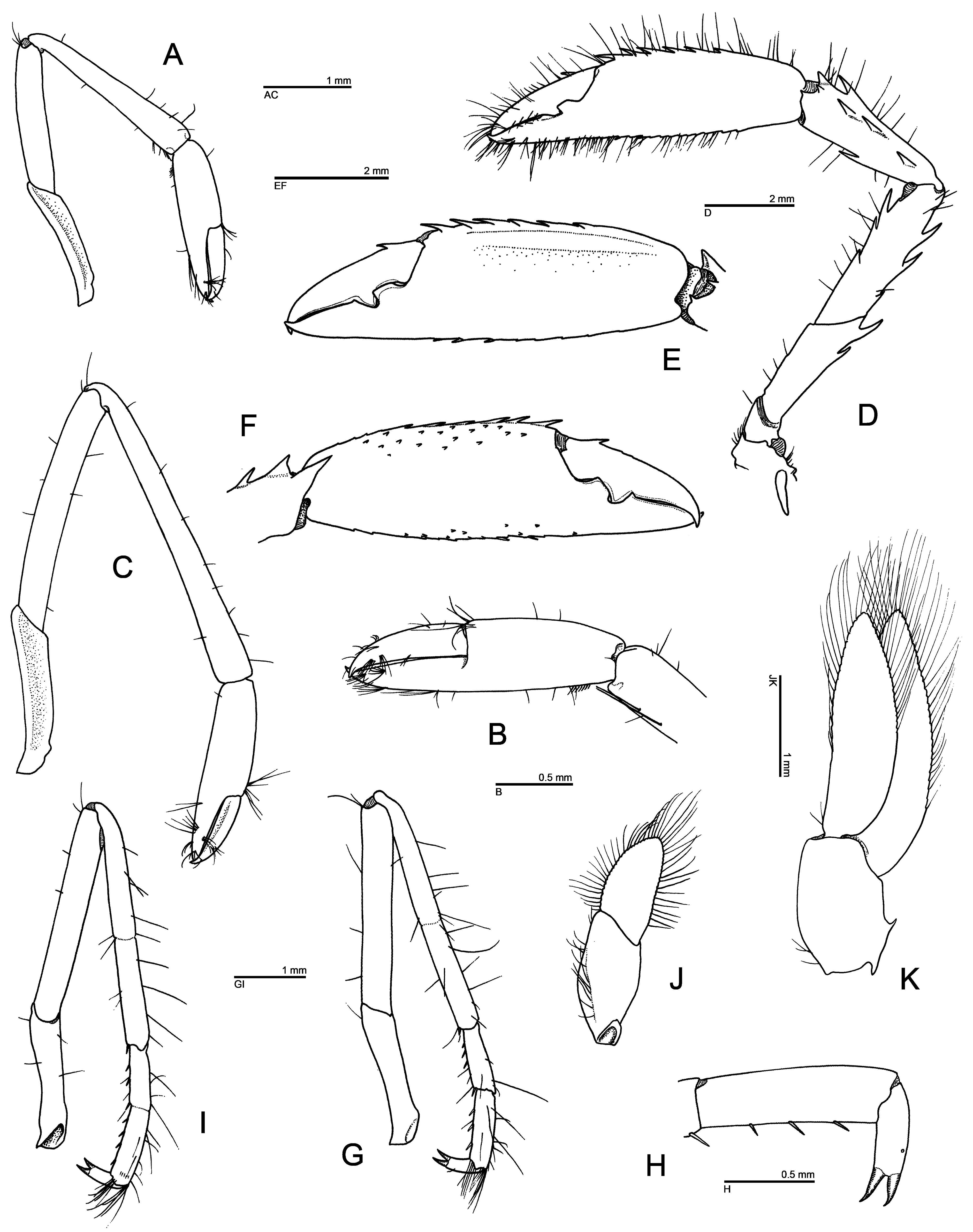
Diagnosis. Medium-sized stenopodidean shrimps with moderately stout, laterally compressed body. Carapace with deep cervical groove and adjacent cervical cincture armed with row of prominent, anteriorly directed spines, some of them larger than antennal spine; post-cervical groove and cincture absent (in J. alucia sp. nov.) or present, armed with prominent spines (in J. foresti ); antennal spine strong; anterolateral margin ventral to antennal spine with several spines, including prominent branchiostegial spine; pterygostomial angle with small spine; supra-orbital and hepatic spines prominent, latter in post-antennal position; remaining surface and margins of carapace unarmed. Rostrum moderately long, compressed, dentate both on dorsal and ventral margins. Pleonites 1–5 unarmed, except for small teeth on posteroventral margin of pleura 2–5 or 4–5; pleonite 6 with posterior margin bluntly protruding, with (in J. foresti ) or without (in J. alucia sp. nov.) small ventrolateral tooth on each side; sternites of pleonites 3–5 each with strong, subacute, median process. Telson narrow, lance-shaped, with narrow median groove flanked by 2 broad, longitudinal carinae, latter unarmed (in J. foresti ) or armed (in J. alucia sp. nov.) with few small spines; lateral margin with prominent spine near mid-length; posterior margin with posterolateral angles protruding as small spines, margin between them unarmed. Eyes somewhat reduced, with globular, pale-pigmented cornea. Dorsal surface of eyestalk with several small spines, some at proximal margin of cornea. Antennular peduncle with small, bluntly ending stylocerite.Antenna with basicerite armed with spines distally; scaphocerite well developed, narrow, with obtusely serrate or smooth lateral margin and two faint longitudinal carinae. Mandible with fused molar and incisor processes, latter with obtuse teeth; palp 3-articulated. Maxillule with broad, distally rounded coxal endite, truncate basial endite and simple, setose endopod (palp). Maxilla with slender, subtly biarticulated endopod (palp); both coxal and basial endites bilobed; scaphognathite slender, its ventral margin with some considerably elongate setae. Maxilliped 1 with 3-articulated endopod (palp); basial endite broad; exopod and epipod well developed, latter bilobed; arthrobranchs absent. Maxilliped 2 with distal endopodal articles distinct; propodus with ventral margin unarmed; exopod well developed; epipod with podobranch; single arthrobranch present. Maxilliped 3 with stout endopod; ischium with (in J. alucia sp. nov.) or without (in J. foresti , to be confirmed) movable spines on ventral margin; merus with spines and sometimes also with movable spines; carpus, propodus and dactylus unarmed, setose; propodus without setiferous organ. Pereiopod 1 with reduced carpo-propodal setiferous organ; chela with simple, unarmed fingers. Pereiopod 2 with all articles unarmed; chela with simple, unarmed fingers. Pereiopods 3 subequal in size, subsymmetrical in shape, much stouter and longer than pereiopods 1 and 2; ischium, merus and carpus armed with few large spines; chela moderately enlarged; palm armed with at least 1 spine, typically row of stronger spines on dorsal margin; dactylus usually armed with 1 or several strong spines on dorsal margin, sometimes unarmed. Pereiopods 4 and 5 rather stout; carpus and propodus each usually subdivided into 2 units; carpus with distoventral movable spinule; propodus with small movable spines along ventral margin; dactylus very stout, strongly biunguiculate. Pleopods biramous except for first pair, without appendices internae; protopods with posterior margin unarmed (in J. foresti ) or armed (in J. alucia sp. nov.) with small subacute tooth. Uropods with lateral margins of exopod and endopod entire (in J. foresti ) or serrated on exopod only (in J. alucia sp. nov.); dorsal surfaces each with 2 blunt, longitudinal carinae, without spines.
Etymology. The genus is named after Dr. Joseph (Joe) Goy, for his significant contribution to the systematics of the Stenopodidea . Gender feminine.
Type species. Jogoya alucia sp. nov., by present designation.
Other species included. Jogoya foresti ( Hendrickx, 2002) , comb. nov. (originally described as Odontozona foresti Hendrickx, 2002 View in CoL ).
Distribution. Tropical western Atlantic ( Bahamas, Anguilla) and eastern Pacific ( Mexico).
Remarks. Jogoya gen. nov. can be separated from each of the morphologically closest genera, viz. Richardina and Odontozona (s. lat., see above), by at least two characters. Jogoya gen. nov. differs most obviously from both Richardina and Odontozona by the unusually strong armature on the posterior margin of the cervical cincture, which consists of 10–11 very large spines on each side of the carapace ( Fig. 1A, B, D View FIGURE 1 ; Hendrickx 2002: fig. 1A, B). In the other two genera, including all mesophotic and deep-water species of Odontozona (see below), the armature of the cervical cincture of the carapace is more modest, consisting of smaller, although sometimes more numerous spines (e.g., Goy 1982: fig. 1B; Dounas & Koukouras 1989: fig. 1; Goy & Cardoso 2014: fig. 1, 4A; Anker & Tavares 2013: fig. 1A; Goy 2015: fig. 24; Criales & Lemaitre 2017: fig. 2A; see also below). The new genus differs from Richardina by the stout and strongly biunguiculate dactyli on the fourth and fifth pereiopods ( Fig. 3H View FIGURE 3 ; Hendrickx 2002: fig. 3d1). In all species of Richardina , the dactyli are slender and simple (e.g., Lo Bianco 1903: pl. 7, fig. 7; Kemp 1910: pl. 23, fig. 1; Goy 1982: fig. 1H; Saito & Komatsu 2009: figs. 1, 4G, H). Furthermore, Jogoya gen. nov. can be separated from most species of Odontozona (= Odontozona s. str.) by the absence of scattered small spines or rows of spines on the carapace posterior to and below cervical (or post-cervical) cincture, and the absence of sculpture on the pleon. The two species of the new genus also show a reduced armature on the dorsal surface of the telson, with only a few spines or no spines at all ( Fig. 1J View FIGURE 1 ; Hendrickx 2002: fig. 1C).
The second species of the new genus, J. foresti , is presently known from the original description ( Hendrickx 2002, as Odontozona foresti ) and two subsequently records ( Hendrickx 2008; Hendrickx & Ayon-Parente 2014, both as O. foresti ), all from the eastern Pacific off Mexico. The eastern Pacific species presents numerous differences in morphology and colour pattern with the western Atlantic type species of the new genus (see below) and therefore its inclusion in Jogoya gen. nov. is somewhat tentative and needs confirmation. In addition, J. foresti seems to have an unusual sexual dimorphism in the armature of the third pereiopod ( Hendrickx & Ayon-Parente 2014) and further specimens are needed to understand the full morphological variation within this species.
The generic position of Odontozona spongicola and several other deep-water or mesophotic species of Odontozona not included in the molecular analysis of Chen et al. (2016) and morphologically clearly different from the herein described new species and O. foresti , viz. O. edwardsi ( Bouvier, 1908) , O. spinosissima Kensley, 1981 , O. striata Goy, 1981 , O. libertae Gore, 1981 , O. minoica Dounas & Koukouras, 1989 , O. joegoyi Hendrickx & Ayon-Parente, 2014 , O. lopheliae Goy & Cardoso, 2014 , and O. spiridonovi Chen & Chan, 2021 (see also Chen & Chan 2021b: table 1) remains to be further investigated. The phylogenetic position of O. spongicola in the same clade as Richardina and Globospongicola , which is very distant from the other, mainly shallow-water and mesophotic species of Odontozona ( Chen et al. 2016: fig. 2) suggests a non-monophyly of Odontozona in its present composition. The position of the mesophotic O. meloi Anker & Tavares, 2013 close to the shallow-water species of Odontozona and the position of the deep-water Juxtastenopus spinulatus ( Holthuis, 1946) among shallow water lineages in Chen et al. (2016: fig. 2), indicate that the two major stenopodidean clades cannot be ecologically defined by depth alone (see also below).
The morphological similarities between two species of Jogoya gen. nov., five species of Richardina and four deep-water species of Odontozona , namely O. spongicola , O. edwardsi , O. joegoyi and O. lopheliae , are obvious. They all share the same type of armature of the carapace, i.e., a cervical cincture with very large (new genus) to moderate or small, continuous spines (vs. more spaced spines in other species of Odontozona ); no spines present in the posterior half of the carapace or below cervical cincture (vs. with scattered spines or rows of spines or spinules in other species of Odontozona ); and the smooth, unsculptured pleon, lacking or with a reduced armature on the pleura and sixth pleonite (vs. sculptured and typically with some spines in other species of Odontozona ) (see also Chen & Chan 2021a). The remaining deep-water / mesophotic species of Odontozona , i.e., O. meloi , O. spinosissima , O. striata , O. libertae , O. minoica and O. spiridonovi , and the 13 presently known shallow-water species, i.e., O. ensifera ( Dana, 1852) [but see below], O. sculpticaudata Holthuis, 1946 (type species of the genus), O. rubra Wicksten, 1982 , O. addaia Pretus, 1990 , O. anaphorae Manning & Chace, 1990 ; O. fasciata Okuno, 2003 , O. crinoidicola Saito & Fujita, 2009 ; O. arbur Saito, Okuno & Anker, 2017 , O. stigmatica Saito, Okuno & Anker, 2017 , O. edyli Criales & Lemaitre, 2017 , O. okunoi Saito & Fujita, 2018 and O. ganzu Saito & Fujita, 2022 (cf. Holthuis 1946; Goy 1981, 2015; Gore 1981; Kensley 1981; Wicksten 1982; Pretus 1990; Manning & Chace 1990; Okuno 2003; Saito & Fujita 2009, 2018, 2022; Anker & Tavares 2013; Goy & Cardoso 2014; Chen & Chan 2021b; Saito et al. 2017; Criales & Lemaitre 2017) together form what appears to be Odontozona s. str., perhaps further subdivided into two groups ( Chen & Chan 2021a). The unusually wide depth range of O. ensifera (2 – 440 m, see Chen & Chan 2021b and references therein) is somewhat suspicious. Thus, the question remains whether the four species of Odontozona presumably allied to O. spongicola and Richardina should be transferred (1) to Jogoya gen. nov., with which they share the biunguiculate dactyli on the fourth and fifth pereiopods; (2) to Richardina , with which they share the relatively moderate spines on the cervical cincture; or (3) to a new genus, which is possibly the most appropriate action (however, beyond the scope of the present study). The paraphyly of Richardina indicated in the rather limited dataset in Chen et al. (2016), as well as the position of Globospongicola , a morphologically highly distinctive, derived lineage, complicate generic assignments in this clade. Including the currently two known species of Globospongicola and four species allied to O. spongicola in Richardina would result in the synonymisation of Globospongicola and a very broadly defined and rather impractical genus Richardina . On the other hand, the reduction of the armature on the carapace, telson and appendages in Globospongicola , as a result of adaptation to symbiotic lifestyle inside hexactinellid sponges ( Komai & Saito 2006; Jiang et al. 2015), is paralleled in some other, presumably non-symbiotic stenopodidean lineages, for instance, Microprosthema lubricum Saito & Okuno, 2011 ( Saito & Okuno 2011).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Stenopodidea |
|
Family |
Jogoya
| Anker, Arthur 2023 |
Jogoya foresti ( Hendrickx, 2002 )
| Anker 2023 |
Odontozona foresti
| Hendrickx 2002 |