
Uca (Petruca) panamensis ( Stimpson, 1859 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4034.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:BBA4A435-BE77-4573-B5C3-2746BEE51A60 |
|
DOI |
https://doi.org/10.5281/zenodo.6113943 |
|
persistent identifier |
https://treatment.plazi.org/id/646187D2-FFD9-706D-1EDC-EB55FB1C763B |
|
treatment provided by |
Plazi |
|
scientific name |
Uca (Petruca) panamensis ( Stimpson, 1859 ) |
| status |
|
Uca (Petruca) panamensis ( Stimpson, 1859) View in CoL
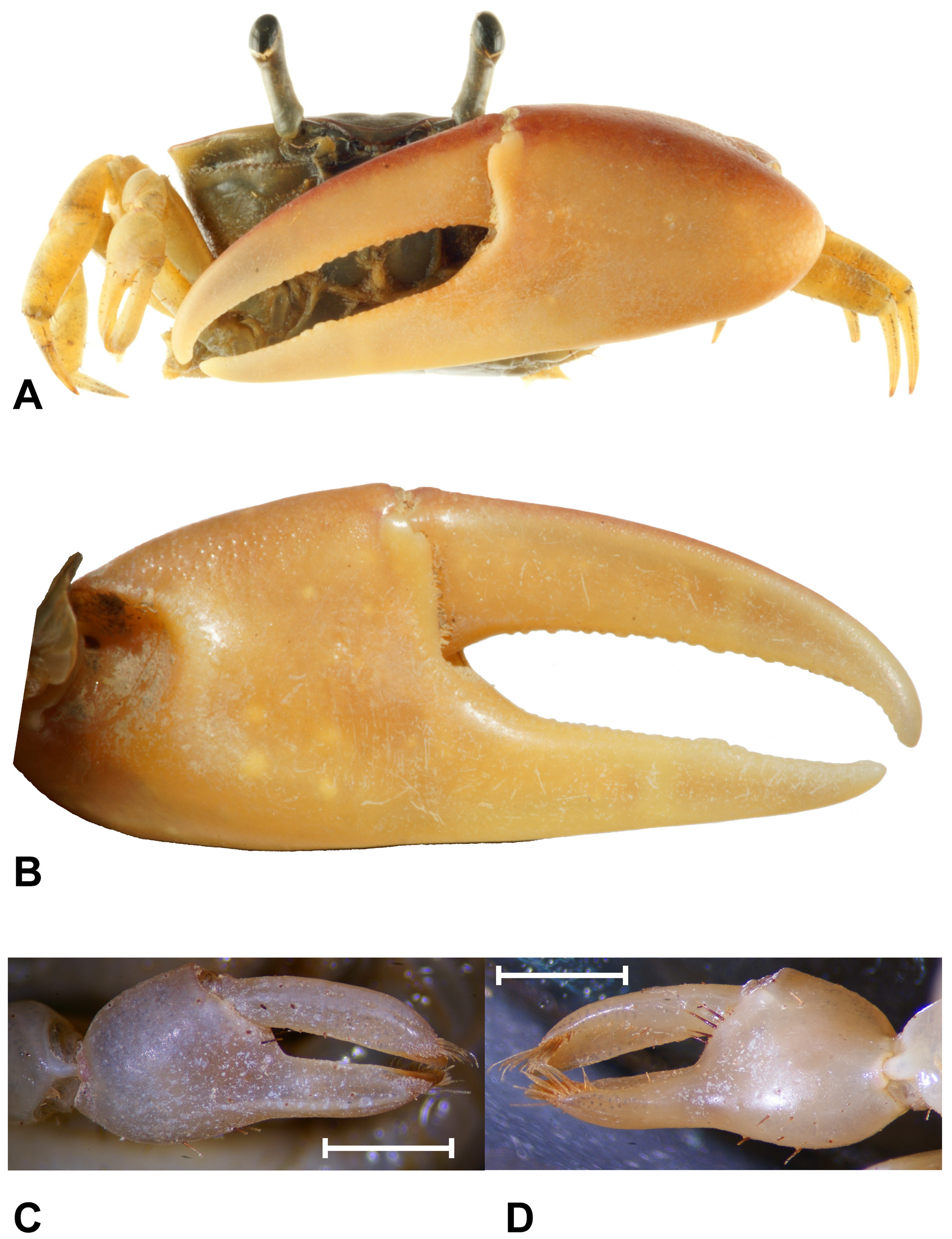
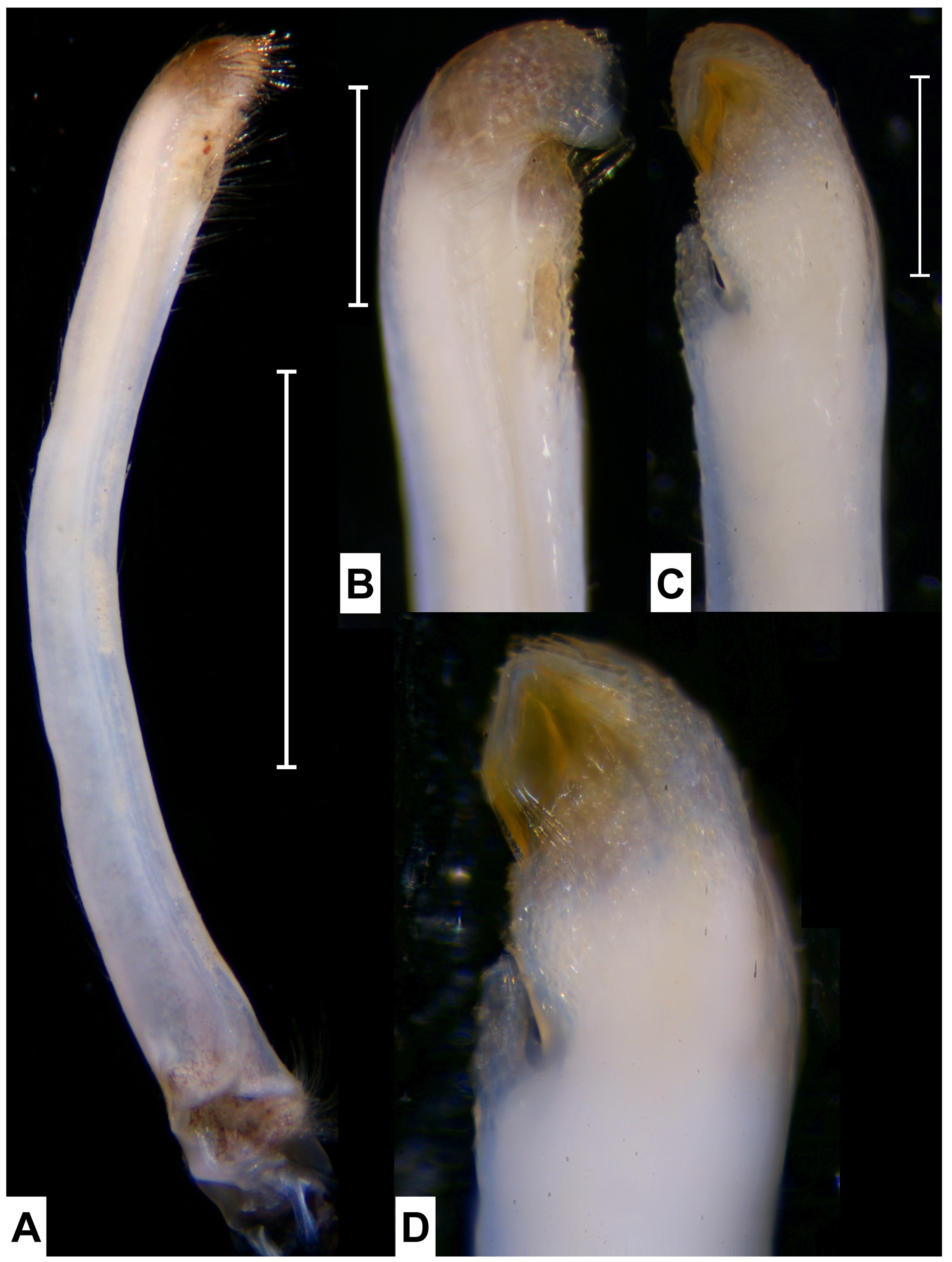
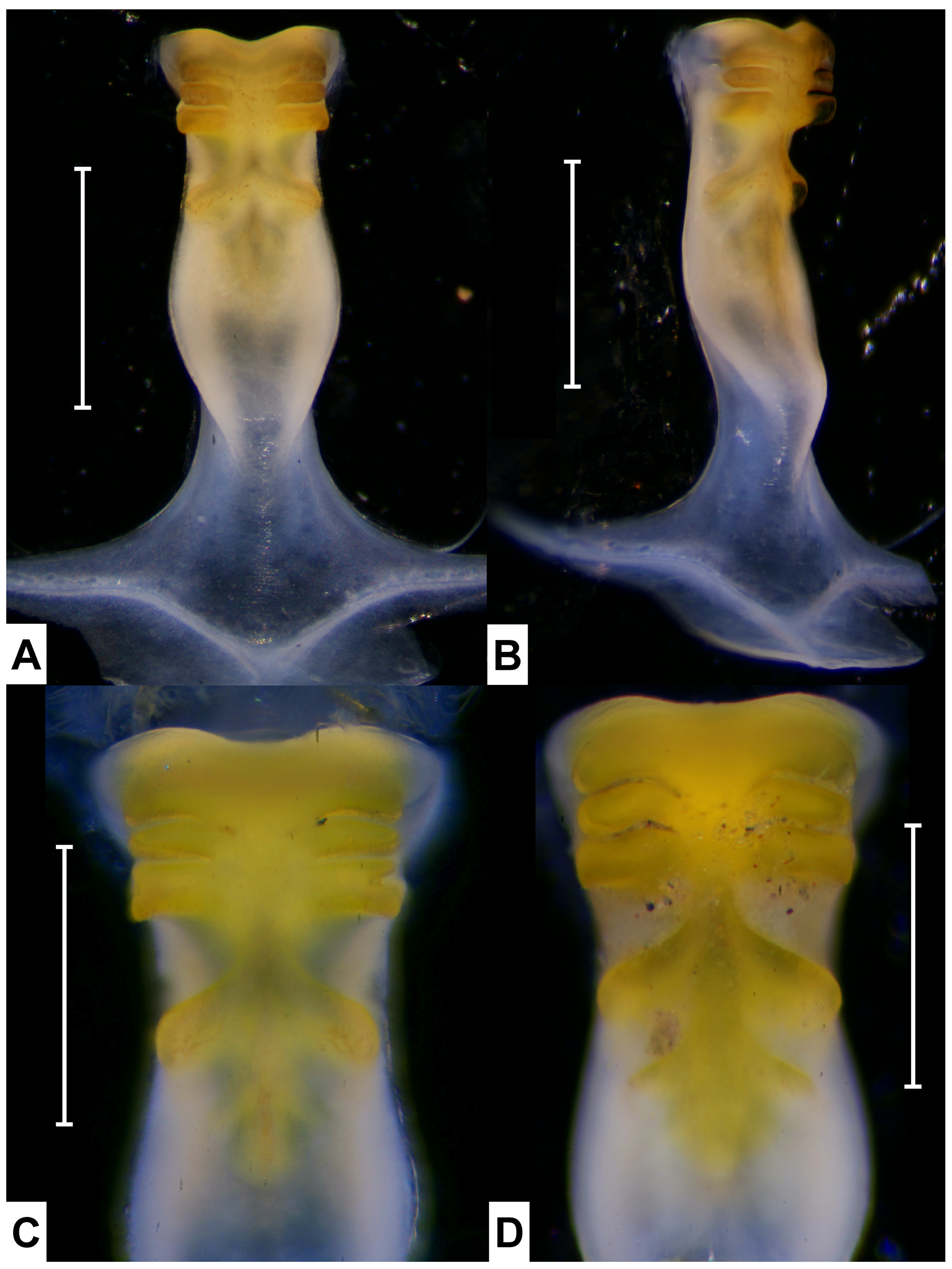
( Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , 8 View FIGURE 8 )
Gelasimus panamensis Stimpson, 1859: 63 View in CoL ; Smith 1870: 139, pl. 4(5); Kingsley 1880: 150, fig. 10(24); Cano 1889: 235.
Uca panamensis View in CoL — Nobili 1901: 49; Rathbun 1918: 412, pl. 149; Crane 1941: 204, test-fig. 4X; Garth 1948: 60; Takeda & Murai 2003: 179, fig. 1.
Uca (Minuca) panamensis View in CoL — Bott 1954: 162, fig. 4, pl. 15(4); Crane 1975: 158, pl. 22A–D, figs. 46H, 66G; Prahl & Guhl 1979: 147, figs. 1–2.
Uca (Leptuca) panamensis View in CoL — Rosenberg 2001: 848, 852; Beinlich & von Hagen 2006: 22, 26, fig. 7j; Ng et al. 2008: 241.
Material examined. Neotype for Gelasimus panamensis , 1 ♂ (16.8 mm), USNM 1294205 (transferred from NCHUZOOL 13581), Culebra Island, Panamá, coll. J. Christy, 2008; 1 ♂ (13.0 mm), NCHUZOOL 13581, same data as neotype; 1 ♂ (12.6 mm), ZRC 2012.0126, Playa San Juanillo, Costa Rica, coll. Z. Jaafar, 25–27 July 2011; 4 ♂♂ (11.1–15.1 mm), 2 ♀♀ (11.7, 13.7 mm), ZRC, 1 ♂ (16.4 mm), 1 ♀ (16.8 mm), NCHUZOOL 14753, San Juanillo rocky shore, Ostional, Costa Rica, coll. Z. Jaafar, 26–29 July, 2011.
Comparative material. See Appendix 1.
Remarks. Gelasimus panamensis was described by Stimpson (1859) from three specimens, all apparently immature, from an undisclosed site in Panamá. No figures were provided. In any case, the types are no longer extant. Like most of his material, they were almost certainly lost in the great Chicago Fire (see Evans 1967; Deiss & Manning 1981; Manning 1993; Vasile et al. 2005; Manning & Reed 2006). Although the species has been well treated by many authors, notably Crane (1975), since a new subgenus is established here for the species, the designation of a neotype for Gelasimus panamensis Stimpson, 1859 , is important for longer term taxonomic stability. This is especially important given that there is now active research on Uca . To this effect, we here select an adult male with CW of 16.8 mm collected from Panamá as the neotype of the species. It is a recently collected specimen (USNM 1294205) that also has genetic data.
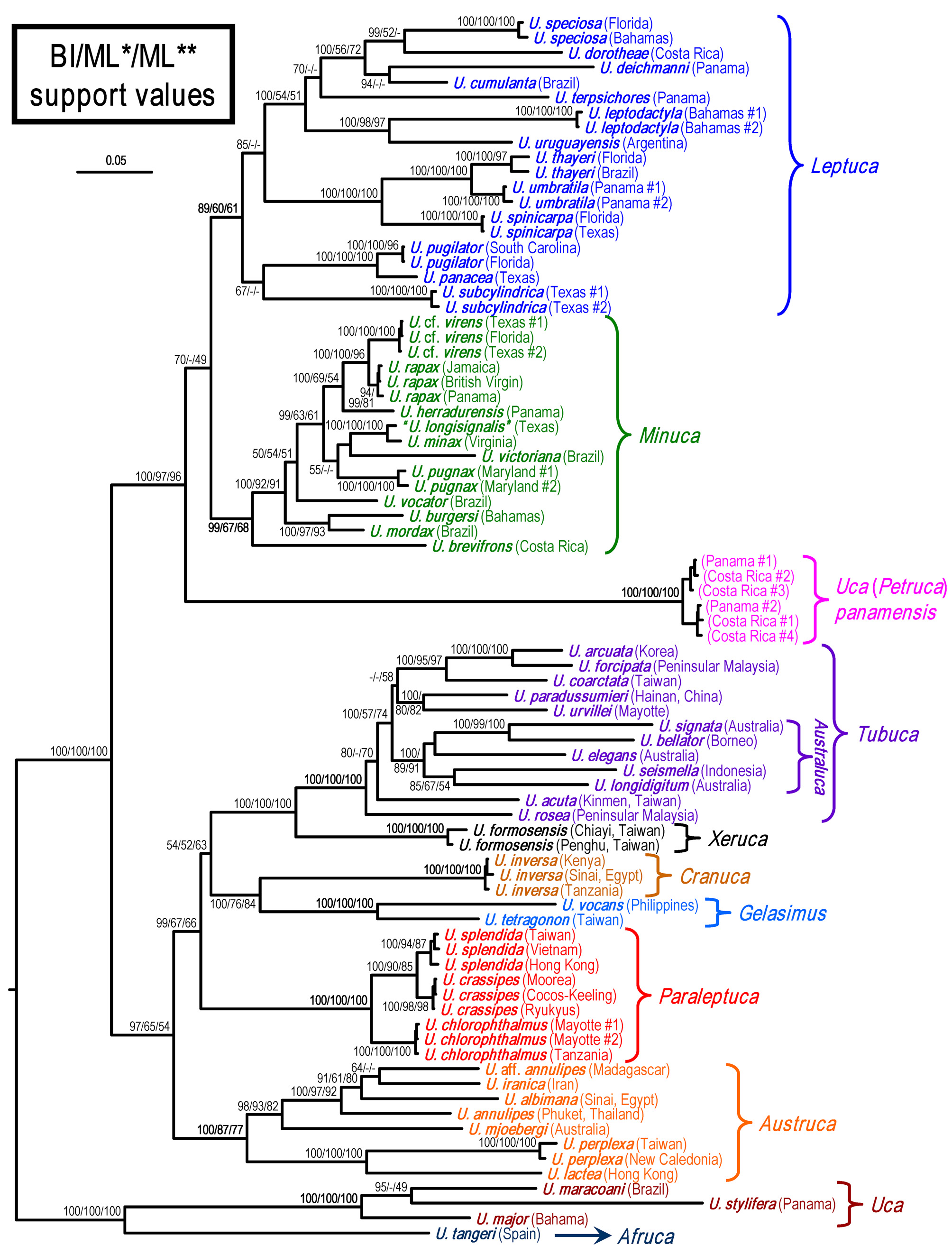
DNA analyses and discussion. A 589 bp segment of the 16S, 658 bp segment of COI and 689 bp segment of 28S, from 55 species of fiddler crabs, were amplified and aligned ( Table 1). The phylogenetic tree of the combined markers was reconstructed from the BI analysis, with the support values from two methods of ML analyses ( Fig. 9 View FIGURE 9. A ). To estimate the divergence time, the substitution rate of 1.17% /10 6 year for the combined 16S and COI of marine sesarmids (cf. Schubart et al. 1998; Ragionieri et al. 2009) was applied to the p-distance between clades.
The relationships of the IWP subgenera ( Fig. 9 View FIGURE 9. A ) generally agree with Shih et al. (2013b) and Shih (2015) and are not repeated here. The American broad-fronted species form a highly-supported main clade by three methods, with three distinct clades corresponding to three subgenera with high support at least by the BI method.
The subgenus Petruca subgen. nov. diverged earlier than the others at about 14 million years ago (mya) (pdistance of the combined 16S and COI = 16.45%). Both Minuca and Leptuca are monophyletic, are closely related and diverged at 9 mya (p-distance = 10.63%).
The six specimens of U. panamensis from the Pacific coasts of Costa Rica and Panamá form a highly supported clade supporting the establishment of Petruca subgen. nov. Its intraspecific K2P divergence COI is average 1.73% (0.33–2.81%), which is lower than U. splendida (1.86%; cf. Shih et al. 2012), but higher than other reported fiddler crabs. For example, the largest intraspecific divergences are 1.23% in U. jocelynae (recalculated from Shih et al. 2010); 0.98% in U. annulipes ( Shih et al. 2009) ; about 0.83% in U. pugnax (cf. Sanford et al. 2006); 0.46% in U. sindensis (recalculated from Shih et al. 2015); about 0.32% in U. maracoani (cf. Wieman et al. 2014); and about 0.13% in U. annulipes (= U. aff. annulipes , see Shih et al. 2013b) from East Africa ( Silva et al. 2010). The high genetic variability within a species may be explained by past historical or present barriers (e.g., ocean currents and salinity) affecting their larval dispersal (see López-Duarte et al. 2011; Shih et al. 2015).
Within the Minuca clade, U. brevifrons diverged earliest at 8 mya (p-distance = 9.27%), and is sister to the remaining species of this subgenus. The habitat of U. brevifrons is unusual. This species lives on the muddy banks of freshwater and brackish streams, sometimes even in wet forest several meters from the nearest stream ( Crane 1941, 1975). Perhaps the simpler form of urocardiac ossicles of gastric mill present in this species ( Fig. 6 View FIGURE 6 A) is an ancestral character. Crane (1975) treated U. brevifrons and U. minax as the same “superspecies [ minax ]” but their close relationship is not supported genetically.
The taxonomy of Uca virens Salmon & Atsaides, 1968 , and U. longisignalis Salmon & Atsaides, 1968 , has been debated since the publication of the description of the two species ( Salmon & Atsaides 1968). By comparing the holotypes of the above two species, von Hagen (1980) confirmed them as synonyms of U. rapax and U. minax , respectively.
The synonymization of U. virens as U. rapax has been followed by later studies (e.g. Barnwell & Thurman 1984; Beinlich & von Hagen 2006). Although there are several studies still using the name of “ Uca virens ” ( Rosenberg 2001; Swanson et al. 2013; Pérez-Mozqueda et al. 2014; del Castillo et al. 2015), the identification of the species in these studies will need to be reconfirmed (C. Thurman, pers. comm.).
Based on the allozyme study of hemolymph amylases ( Barnwell & Thurman 1984) and esterase-1 and -2 ( Salmon & Kettler 1987), it is apparent that there are two “forms” of U. rapax , corresponding to the distribution of U. virens and U. rapax . In our study based on 16S, COI and 28S markers ( Fig. 9 View FIGURE 9. A ), there is a clade sister to U. rapax with similar morphology. Before additional studies are done to confirm whether it is actually U. virens , we use the name “ U. cf. virens ” for convenience. The two species diverged at 2 mya (p-distance = 2.32%), although Salmon & Kettler (1987) estimated the divergence time is 20–50 thousand years based on the allozyme data.
While previous studies considered the morphological characters of U. virens were within the range of U. rapax ( Hagen 1980; Barnwell & Thurman 1984), their gastric mill structures show differences in the posterior tooth plate of urocardiac ossicles, with 8 ridges in U. rapax ( Fig. 6 View FIGURE 6 F) and 6 ridges in U. virens ( Fig. 6 View FIGURE 6 B). More studies of morphological characters, especially the gastric mills, are necessary to clarify the identity of U. virens .
Another confused species is U. longisignalis , which has been synonymized as U. minax by von Hagen (1980). Thurman (1982) considered it is a valid species because there are substantial morphological differences between the two species, a decision that has been followed by other studies thereafter. Genetically, Felder & Saton (1994) showed that the allozyme evidence did not support the hypothesis that U. longisignalis is different from U. minax . Landstorfer et al. (2008) and Warwick et al. (2009) also reported the failure to separate the two species by mitochondrial COI or nuclear internal transcribed spacer (ITS-1) markers. We included in our study specimens of the two species with typical characters of each taxon (mainly based on with or without pubescence ventrally; cf. Barnwell & Thurman 1984), but the molecular data only shows one clade ( Fig. 9 View FIGURE 9. A ), therefore agreeing with earlier genetic studies. Further studies, with more complete sampling, will be necessary to clarify the taxonomy of the U. minax complex.
The close relationship of U. burgersi and U. mordax suggested by Salmon & Kettler (1987) is highly supported ( Fig. 9 View FIGURE 9. A ). Within the Leptuca clade, U. panacea , and U. pugilator are closely related, with the divergence time at 2.8 mya (p-distance = 3.24%), which supports the original hypothesis that both species have diverged recently ( Novak & Salmon 1974; Barnwell & Thurman 1984).
The supposed close relationship of U. speciosa and U. spinicarpa ( Crane 1975; Barnwell & Thurman 1984) is not supported in this study ( Fig. 9 View FIGURE 9. A ). Our study, however, agrees well with the allozyme studies ( Salmon et al. 1979; Salmon & Kettler 1987) that the genetic distance of two species is large. Uca speciosa is distinct from other species genetically, although it forms a main clade, with U. dorotheae , U. deichmanni , and U. cumulanta . Uca spinicarpa is, however, closely related to the species pair, U. thayeri and U. umbratila (see below).
Crane’s (1975) subgenus Boboruca (= Planuca Bott, 1973 ), established for U. thayeri and U. umbratila , was synonymized with Minuca by Albrecht & von Hagen (1981), based on combined allozyme and morphological characters, and this has been followed by all later authors (e.g., von Hagen 1987; Salmon & Zucker 1988; Rosenberg 2001; Beinlich & von Hagen 2006; Ng et al. 2008). In this study, U. thayeri and U. umbratila came out clearly inside the Leptuca clade ( Fig. 9 View FIGURE 9. A ). In the studies of Levinton et al. (1996) and Sturmbauer et al. (1996) using the 16S marker, the position of U. umbratila is within a “derived American clade” composed of a mixture of the subgenera Minuca and Leptuca . Uca umbratila is nevertheless located within Minuca in Landstorfer & Schubart's (2010) study, although their sequences of COI ( FN430708 View Materials ) and 28S ( FN430719 View Materials ) agree well with U. umbratila in our study. The different conclusions are probably caused by the inclusion of 16S marker, as well as the number of species used in ingroups and outgroups. Hampton et al. (2014) have pointed out the unsolved relationship of Boboruca from the above studies. Our study gives genetic support for including U. thayeri and U. umbratila in the subgenus Leptuca ( Fig. 9 View FIGURE 9. A ), although their morphology is closer to Minuca .
One of the main characters placing these two species in Minuca is the presence of two posterolateral striae on each side of carapace (cf. Beinlich & von Hagen 2006; Bezerra 2012). This character, however, can be variable. While there are two striae on each side of the carapace in U. thayeri , U. umbratila only has one stria (cf. Crane 1941: text-fig. 7). This has not been discussed before. The available data also implies the morphological similarity between the two species and Minuca is convergent.
The present study shows that U. thayeri , U. umbratila , and U. spinicarpa form a highly supported clade which may be the result of allopatric speciation. Whereas U. thayeri and U. umbratila diverged at 3.7 mya (p-distance = 4.34%), a well-known species pair separated after the closure of the Isthmus of Panamá around 3.1 million years ago (reviewed by Schubart et al. 1998; Lessios 2008), U. spinicarpa is endemic to the Gulf of Mexico in a more northern distribution, and it diverged from the other two species at 6.3 mya (p-distance = 7.38%). Uca subcylindrica is endemic along the border between Texas and northern Mexico in the Gulf of Mexico, and has been considered as an anomalous form, with unusually rounded body, striking form of G1 and hypersaline lagoonal habitat, without close alliance to other species ( Crane 1975; Barnwell & Thurman 1984; Thurman 1987). Crane (1975) placed it in Minuca , but noted with reservation that it was close to Leptuca . Because some characters of U. subcylindrica are shared with several members of Leptuca, Barnwell & Thurman (1984) considered it was not suitable to retain in the homogeneous Minuca and transferred it to Leptuca . Their action is supported by our study, as U. subcylindrica forms a distinct clade from other species of this subgenus ( Fig. 9 View FIGURE 9. A ). The simpler form of urocardiac ossicles of the gastric mill ( Fig. 7 View FIGURE 7 A) may also be a reflection of the early split of this species from others.
Uca panamensis View in CoL was placed in Minuca View in CoL mainly by the characters of the front of the carapace and male first gonopod ( Bott 1954, 1973; Crane 1975). Based on a morphological analysis, Rosenberg (2001) placed it in Leptuca View in CoL along with U. terpsichores View in CoL , U. deichmanni View in CoL , U. leptodactyla View in CoL , and U. dorotheae View in CoL . The many unusual features of U. panamensis View in CoL (e.g., flatter carapace, stiff setal brushes on the tips of the small chelae, the posterior extension of major View in CoL manus, preference for rocky habitats etc.) suggest this classification is incorrect. The present use of new morphological characters (viz. the orbital armature at the inner corner of orbital floor, simple form of urocardiac ossicles of gastric mill, large and bent inner process of the G1) as well as molecular evidence using mitochondrial and nuclear markers ( Fig. 9 View FIGURE 9. A ), support the hypothesis that U. panamensis View in CoL belongs to its own subgenus (Petruca subgen. nov.), and diverged from other American Minuca View in CoL and Leptuca View in CoL at about 14 mya.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Uca (Petruca) panamensis ( Stimpson, 1859 )
| Shih, Hsi-Te, Ng, Peter K. L. & Christy, John H. 2015 |
Uca (Leptuca) panamensis
| Hagen 2006: 22 |
| Rosenberg 2001: 848 |
Uca (Minuca) panamensis
| Prahl 1979: 147 |
| Crane 1975: 158 |
| Bott 1954: 162 |
Uca panamensis
| Takeda 2003: 179 |
| Garth 1948: 60 |
| Crane 1941: 204 |
| Rathbun 1918: 412 |
| Nobili 1901: 49 |
Gelasimus panamensis
| Cano 1889: 235 |
| Kingsley 1880: 150 |
| Smith 1870: 139 |
| Stimpson 1859: 63 |