
Calyptotheca conica Cook, 1965
|
publication ID |
https://doi.org/10.11646/zootaxa.3827.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:D9AEB652-345E-4BB2-8CBD-A3FB4F92C733 |
|
DOI |
https://doi.org/10.5281/zenodo.6126947 |
|
persistent identifier |
https://treatment.plazi.org/id/64480F01-FFF9-277D-FF04-B247FC6EFAB8 |
|
treatment provided by |
Plazi |
|
scientific name |
Calyptotheca conica Cook, 1965 |
| status |
|
Calyptotheca conica Cook, 1965 View in CoL
( Figures 3 View FIGURE 3 , 4 View FIGURE 4 ; Tables 3 View TABLE 3 , 4 View TABLE 4 )
Calyptotheca conica Cook, 1965: 437 View in CoL , pl.1, figs 1–3.
Material examined. Holotype: NHMUK 1892.1.28.76A, Holothuria Bank , off Cape Talbot, northwestern Australia, 15–32 fathoms, coll. Basset-Smith. Paratype: NHMUK 1892.1.28.76C, same data as holotype.
Holothuria Bank, Kimberley , north-western Australia: NHMUK 1892.1.28.76G, same data as holotype.
Bryomol Reef, GoC: MTQ G26472, G26509 (SEM stub), 16°31.064' S, 139°56.755' E, 33 m.
Beagle Gulf, Northern Territory: MAGNT G228, Stevens Rock, Darwin Harbour, 12.48556667° S, 130.78515° E, 8 November 2004, pebble in sand, 12 m, coll. K. Gowlett-Holmes; MAGNT G27, Fish Reef, Bynoe Harbour, 12.414° S, 130.482° E, 6 October 1993, coarse sand and gravel, 17 m, coll. S.K. Horner; MAGNT G22, Outer Darwin Harbour, 12.168° S, 130.782° E, 12 October 1993, mud and gravel, 30 m, coll. R.S. Williams; MAGNT G5, Middle of Shoal Bay, 12.218° S, 130.933° E, 12 October 1993, sandy mud and sparse seagrass, 17 m, coll. R.S. Williams.
Description of holotype. Colony a unilaminar, hollow cone, basal surface with rigid, hollow, calcareous hooks. Autozooids rectangular to hexagonal, longer than wide (c. 0.4 × 0.3 mm, Table 3 View TABLE 3 ). Frontal shield flattened to slightly convex, with regularly spaced, rounded pseudopores (average 42 per zooid), each in a pit, separated by more than the width of a pseudopore, extending to zooid borders, absent proximal to orifice. Interzooidal boundaries marked by thin, raised, sinuous suture lines that curve around marginal avicularia.
Primary orifice rounded, wider than long (c. 0.09 × 0.10 mm), lunula extending to lateral edges of orifice, poster with wide, shallow sinus, condyles large, bilobed, knuckle-shaped, non serrate, ridged proximally. No suboral umbo.
Adventitious avicularia small, oval to rounded-subtriangular, slightly longer than wide (c. 0.06 × 0.05 mm), marginal (borne laterally against interzooidal suture lines), oriented to centre of frontal shield; also occurring in each distal angle and up to several more medio- or proximolaterally, or proximally, opesia oval proximally, semicircular distally, crossbar complete.
Vicarious avicularia uncommon, longer than autozooids (one measured c. 0.7 × 0.4 mm); frontal shield with regularly spaced, rounded pseudopores, similar in appearance, size and spacing to those of autozooids, extending to lateral margins, absent proximal to orifice. Avicularian orifice much larger than in autozooids, rounded and slightly wider distally (one measured c. 0.5 × 0.3 mm), lateral margins straight, slightly converging proximally, a shallow sinus proximally; condyles large, not bilobed or serrate, suboral umbo absent. Marginal adventitious avicularia present, similar in size, shape and orientation to those of autozooids.
Ovicell immersed, not raised above surrounding zooids, with pseudopores of similar size and density to frontal shield, secondary calcification cormidial as indicated by Y-shaped suture lines. Orifice dimorphic, wider than in autozooid (c. 0.13 vs 0.10 mm; Table 3 View TABLE 3 ).
Variations in GoC and Beagle Bay specimens. The GoC specimen has larger orifices than the holotype (c. 0.11 × 0.12 mm vs 0.09 × 0.10 mm for autozooids; c. 0.11 × 0.16 mm vs 0.09 × 0.13 mm for ovicellate zooids; Table 4 View TABLE 4 ), though with similar dimensions, sinus and condyles. Adventitious avicularia are on average slightly smaller than those of the holotype (c. 0.05 × 0.04 mm vs 0.06 × 0.05 mm). The vicarious avicularia of the GoC specimen are slightly smaller than the single one measured in the holotype (c. 0.6 × 0.4 mm vs 0.7 × 0.4 mm), and their orifices, though similar in shape (elongate-oval), are much smaller, occupying less than half the length of the avicularium vs approximately three-quarters the length in the holotype (0.2 × 0.2 mm vs 0.5 × 0.3 mm).
One of Cook’s (1965) seven Calyptotheca conica specimens (76G) differed sufficiently from the others to not be included as a paratype. It resembles the GoC specimen in the much smaller orifices of the vicarious avicularia (c. 0.2 × 0.1 mm; Cook 1965). Colony 76G is a tiny colony, possibly younger, with none of the frontally budded areas present in the holotype and paratypes.
Cook (1965) reported the ratio of vicarious avicularia to autozooids as 1:82 for the holotype and paratypes and 1: 16 in colony 76G. We recorded 1:17 for Beagle Gulf specimen MAGNT G5 and 1:76 for the GoC specimen (MTQ G26472).
The ovicells of the holotype are immersed and crossed by Y-shaped suture lines [not depicted in Cook’s (1965, fig. 1A) illustration]. The ovicells of the GoC specimen are immersed, the depression reaching or approaching the basal lamina of the distal autozooid. They differ from the holotype in being more prominent and slightly raised above the surrounding zooids, and having larger pseudopores than the frontal shields ( Fig. 4 View FIGURE 4 C), but these features likely change with further secondary calcification (the zooids in Fig. 4 View FIGURE 4 C are at the growing edge of the colony).
Remarks. Calyptotheca conica is distinguished by its large, bilobed condyles, abundant marginal adventitious avicularia, large vicarious avicularia (which also have marginal adventitious avicularia), and a unilaminar encrusting or conical growth form with hooked processes on the basal surface. It resembles C. wasinensis in the shape of the orifice, in having numerous oval marginal avicularia of similar size, and dimorphic orifices. In C. conica , however, the orifices are smaller, the condyles are strikingly different, being large and bilobed, and there is no suboral umbo. The GoC specimens of C. wasinensis have marginal avicularia in all four distal and proximal angles, whereas in C. conica they occur in one or both distal angles and most often in mediolateral positions rather than the proximal angles. Both species begin as encrusting colonies and are most readily confused at this stage, but in later ontogeny C. conica may become a wide, hollow cone whereas C. wasinensis forms bilaminar, foliaceous sheets.
Cook (1965), Harmer (1957, p. 1009) and Hastings (1932, p. 422) described the conical growth of specimens from the Holothuria Bank (northwestern Australia) as a possible adaptation to unstable sandy or muddy sea bottoms, resulting from settlement on small objects such as shell fragments that may be the only available substrata, with subsequent growth overhanging the object. Thus, the conical growth form may be substratumdependent and may not occur in all colonies ( Cook 1965). The single GoC specimen of C. conica encrusts the convex side of a broken bivalve shell c. 2.0 × 1.5 cm, but does not overhang the edges of the shell. One specimen from Beagle Gulf (MAGNT G5) encrusts the convex side of a bivalve shell and overhangs it. Another (MAGNT G27) is roughly conical (c. 2.0 cm diameter × 0.8 cm high), with the centre attached to a small pebble. The other Northern Territory specimens (MAGNT G22 and MAGNT G228) are fragments and are not encrusting or attached to a substratum.
Three other Calyptotheca species with conical, free-lying ( sensu Cook 1965) colonies were described by Harmer (1957): C. parcimunita Harmer, 1957 , C. circularis Harmer, 1957 and C. orbiculata Harmer, 1957 . Calyptotheca parcimunita and C. circularis are readily distinguished from C. conica by their lack of vicarious avicularia and in having adventitious avicularia that differ in both shape and position from those of C. conica . In C. parcimunita they are distolateral to the orifice, minute, acute and rare, and in C. circularis they are suboral, paired, acute, and directed medially. It is clear that the presence of vicarious avicularia and basal calcareous hooked processes in C. conica are not essential to the conical form or to life in an unstable habitat, as C. parcimunita , which co-occurs with C. conica in northwestern Australia ( Cook 1965), does not have either.
Calyptotheca orbiculata View in CoL has vicarious avicularia, calcareous projections from the basal walls and immersed ovicells ( Harmer (1957, pp. 1010, 1014). Cook (1965, p. 436) observed a reduced polypide inside a vicarious avicularium. Calyptotheca orbiculata View in CoL differs from C. conica View in CoL in lacking adventitious avicularia.
Vicarious avicularia also occur in non-conical Calyptotheca View in CoL : C. capitifera ( Canu & Bassler, 1929, p. 300), C. inclusa ( Thornely, 1906) View in CoL , C. obscura Harmelin, López View in CoL de la Cuadra & García-Gómez, 1989 and C. reniformis Tilbrook, 2006 View in CoL (K. Tilbrook, unpubl. data). These are all encrusting species, which in theory could grow a conical form, however they all differ from C. conica View in CoL in aspects of avicularia, sinus, condyles and/or lunula. The vicarious avicularium of C. capitifera is spatulate. Harmer (1957, p. 1010) recorded C. capitifera from the Philippines and Indonesia with illustrations of the vicarious avicularia (pl. 68, figs 1–3) that differed from the spatulate form of those in the original figures of Canu & Bassler (1929, pl. 32, fig. 5), suggesting that Harmer’s specimens (1957) are not C. capitifera sensu stricto. Both C. inclusa View in CoL and C. reniformis View in CoL have a single, suboral adventitious avicularium, a deep U-shaped sinus and serrate condyles. C. obscura View in CoL is an Eastern Atlantic/Mediterranean species with lateral, acute adventitious avicularia, a deep triangular sinus and serrate condyles.
Cook (1965) reported the presence of a polypide in the vicarious avicularia of Calyptotheca conica View in CoL but the presence or absence of polypides could not be confirmed in the present material. The vicarious avicularium of C. obscura View in CoL had a polypide with a reduced number of tentacles (17–18 vs 19–21) and more highly developed opercular muscles compared with autozooids ( Harmelin et al. 1989). Both the reduced polypide and increased musculature are widely accepted as features of primitive avicularia, like the B-zooids of certain Steginoporella View in CoL species, proposed by Harmer (1900), Banta (1973) and others to have evolved from autozooids.
Hirose (2010, p. 117, pl. 201) described Calyptotheca wasinensis View in CoL from Sagami Bay, Japan, as encrusting, with immersed ovicells and vicarious avicularia (the condyles were not described). These characters resemble those of C. conica View in CoL but the autozooidal orifice is larger and more oval (c. 0.15 × 0.19 mm vs 0.09 × 0.10 mm for C. conica View in CoL ), suggesting he had a separate species, which would raise the number of known Calyptotheca View in CoL species with vicarious avicularia to eight.
Distribution. Calyptotheca conica was originally described from Holothuria Bank off Cape Talbot, far north Western Australia. Its distribution is here extended eastwards to Beagle Gulf and Darwin Harbour, Northern Territory, and Bryomol Reef, southeastern GoC, where specimens were collected from sand, gravel and mud substrata at 12– 33 m. A single specimen was found at Bryomol Reef at 33 m.
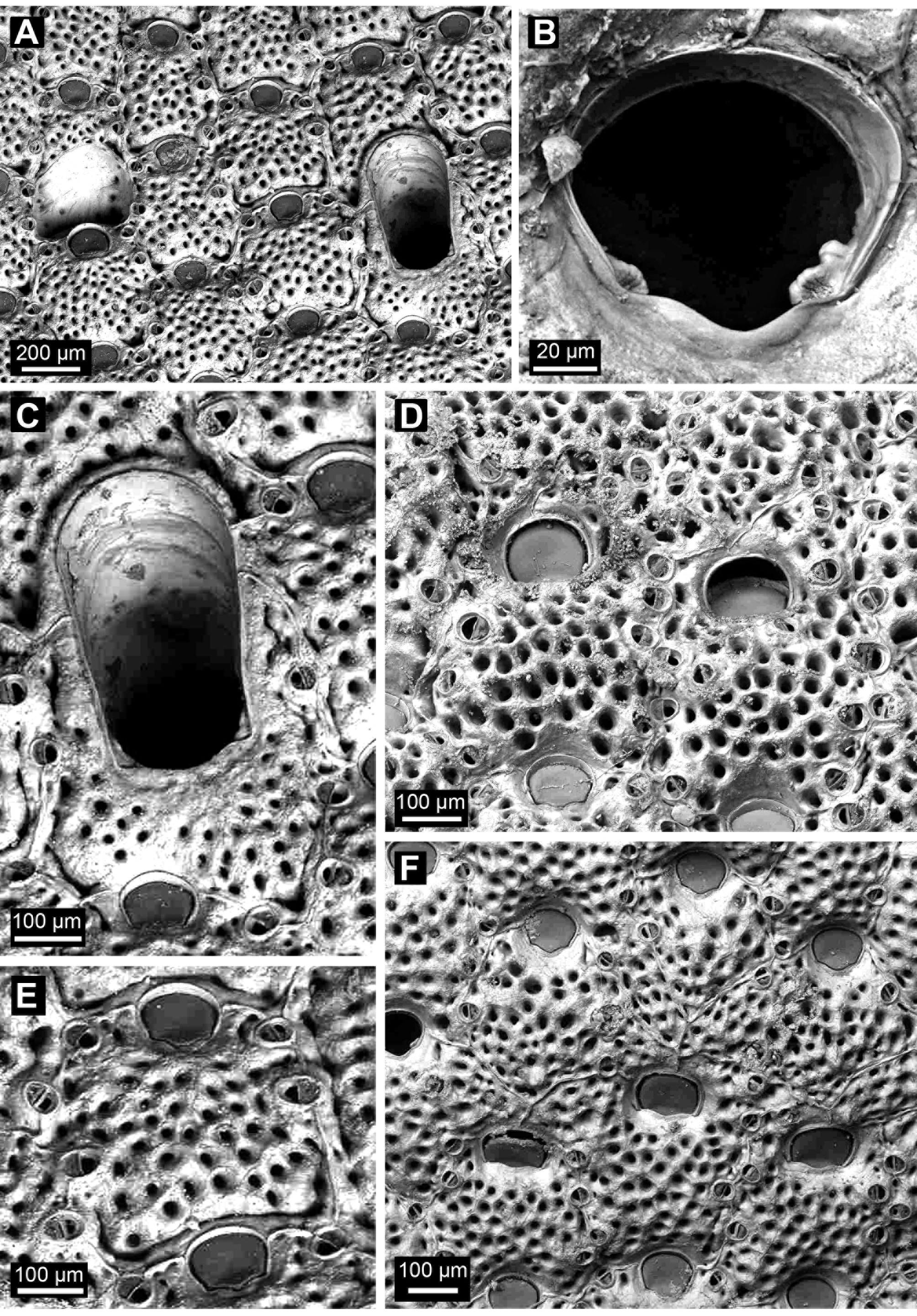
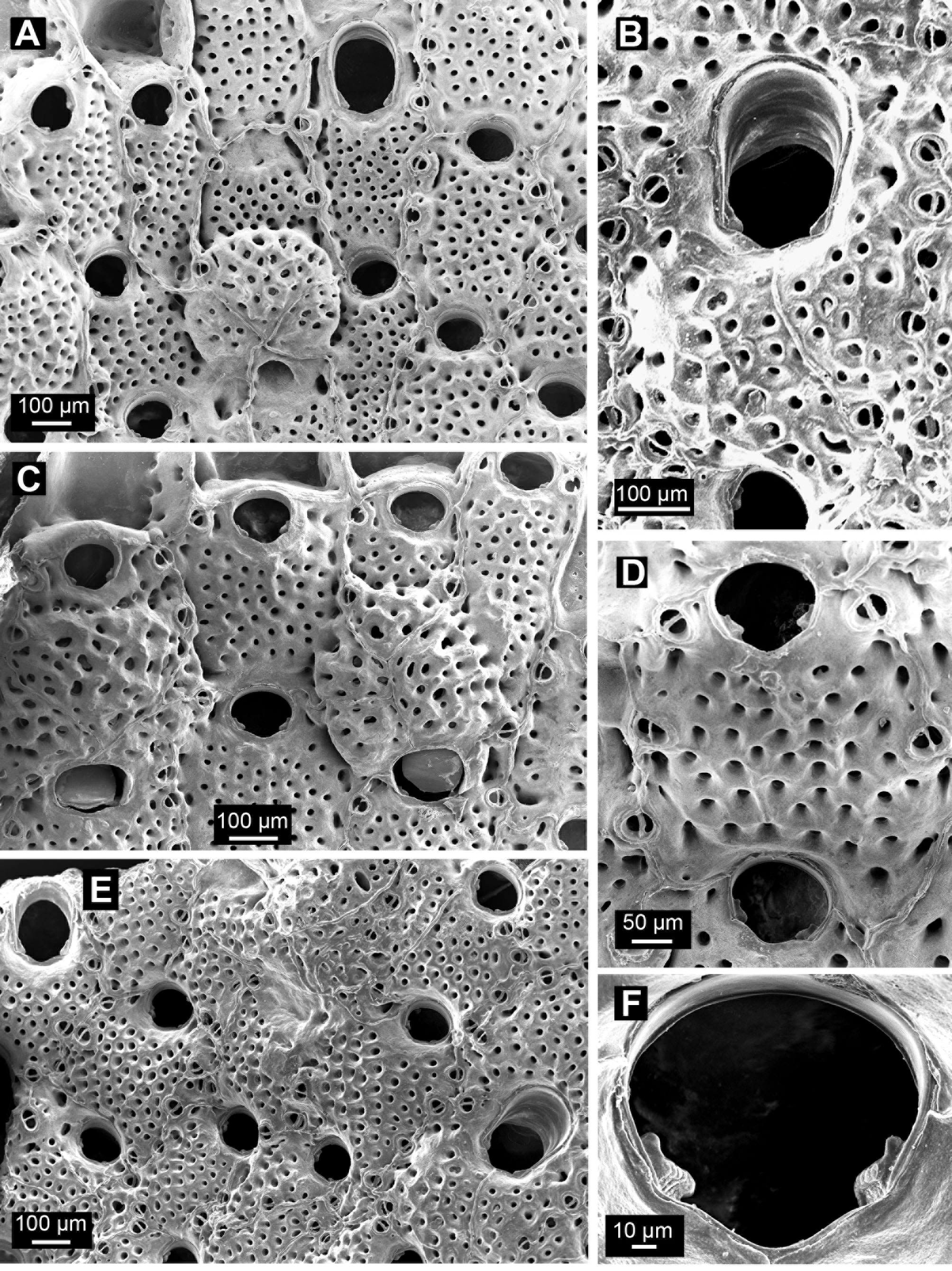
TABLE 3. Measurement (mm) and count data for Calyptotheca conica holotype (NHMUK 1892.1.28.76 A).
| Zooid length | Mean 0.400 | Standard Deviation 0.023 | Range N 0.364–0.444 20 |
|---|---|---|---|
| Zooid width Autozooid orifice length Autozooid orifice width | 0.325 0.0852 0.1048 | 0.051 0.006 0.005 | 0.266–0.410 20 0.078–0.100 20 0.098–0.114 20 |
| Adventitious avicularium length Adventitious avicularium width Vicarious avicularium length | 0.061 0.047 0.692 | 0.007 0.003 | 0.047–0.074 20 0.042–0.052 20 1 |
| Vicarious avicularium width Vicarious avicularium orifice length Vicarious avicularium orifice width | 0.431 0.471 0.273 | 1 1 1 | |
| Diameter of perforations in frontal shield Number of perforations in frontal shield Ovicellate orifice length | 0.015 41.90 0.092 | 0.001 4.56 0.012 | 0.014–0.018 10 37–51 10 0.080–0.105 4 |
| Ovicellate orifice width | 0.134 | 0.004 | 0.131–0.139 4 |
TABLE 4. Measurement (mm) and count data for Calyptotheca conica from the GoC (all measurements on one colony: MTQ G 26472, MTQ G 26509 [SEM stubs]).
| Zooid length | Mean 0.416 | Standard Deviation 0.050 | Range N 0.367–0.531 10 |
|---|---|---|---|
| Zooid width Autozooid orifice length Autozooid orifice width | 0.332 0.109 0.124 | 0.081 0.006 0.004 | 0.207–0.470 10 0.100–0.115 10 0.119–0.130 10 |
| Adventitious avicularium length Adventitious avicularium width Vicarious avicularium length | 0.051 0.042 0.580 | 0.008 0.004 0.011 | 0.040–0.064 10 0.038–0.050 10 0.571–0.588 2 |
| Vicarious avicularium width Vicarious avicularium orifice length Vicarious avicularium orifice width | 0.363 0.240 0.168 | 0.083 0.022 0.011 | 0.304–0.422 2 0.215–0.259 3 0.160–0.181 3 |
| Diameter of perforations in frontal shield Number of perforations in frontal shield Ovicell length | 0.014 46 0.328 | 0.002 7.62 0.013 | 0.011–0.017 10 34–58 8 0.314–0.342 4 |
| Ovicell width Ovicellate orifice length Ovicellate orifice width | 0.387 0.106 0.157 | 0.041 0.004 0.001 | 0.353–0.439 4 0.101–0.109 3 0.156–0.159 3 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Neocheilostomina |
|
Family |
|
|
Genus |
Calyptotheca conica Cook, 1965
| Cumming, Robyn L. & Tilbrook, Kevin J. 2014 |
Calyptotheca conica
| Cook 1965: 437 |