
Citipati osmolskae Clark et al., 2001
|
publication ID |
https://doi.org/10.1206/0003-0082(2002)364<0001:CAOCOT>2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5604437 |
|
persistent identifier |
https://treatment.plazi.org/id/64443907-284A-7E67-FD5E-9DBCFCA7AC13 |
|
treatment provided by |
Felipe |
|
scientific name |
Citipati osmolskae Clark et al., 2001 |
| status |
|
Citipati osmolskae Clark et al., 2001
HOLOTYPE: IGM 100 View Materials /978, a nearly complete skeleton.
TYPE LOCALITY: Djadokhta Formation at Ankylosaur Flats, Ukhaa Tolgod, Gurvan Tes Somon, Omnogov Aimak, Mongolia. Precise coordinates will be made available to qualified researchers on request.
REFERRED SPECIMENS: IGM 100/979, a partial skeleton overlying a nest, from Ankylosaur Flats, Ukhaa Tolgod; IGM 100/971, an embryonic skeleton within an egg.
DIAGNOSIS: Oviraptorid differing from Conchoraptor in having a taller and more highly pneumatized nasal, from Ingenia in that metacarpal I is not extremely broad, from Oviraptor philoceratops in having a shorter skull and mandible, from O. mongoliensis in lacking a convex crest on the frontals and parietals, and from Khaan mckennai in the features listed below. Differs from all other oviraptorids where known in the anterodorsally sloping occiput and quadrate, the parietal being much longer along the midline than the frontal and reaching nearly to the level of the anterior end of the orbit, the ascending process of the jugal being perpendicular to the horizontal ramus (rather than extending posterodorsally), the narial opening being more nearly circular and the ascending process of the premaxilla being vertical rather than sloping posterodorsally, and the cervical vertebrae being elongate (approximately twice as long as they are wide).
REMARKS: This specimen was identified preliminarily as O. philoceratops (e.g., Webster, 1996), but comparison with the holotype of the latter revealed several significant differences (see below). A postcranial skeleton that is preserved on top of a nest, IGM 100/ 979, is referred to the new species and was described by Clark et al. (1999). IGM 100/ 979 is referred to C. osmolskae , pending preparation of the postcranial skeleton of the holotype, primarily on the basis of its large size and its differences from the skeleton of Khaan mckennai . An embryonic skeleton referred to this species was given a preliminary description by Norell et al. (1994) and a more detailed description by Norell et al. (2001). This specimen is referred to Citipati on the basis of its premaxilla, which is nearly vertical rather than sloping posteriorly as in Khaan . An oviraptorid collected from the Djadokhta Formation at Dzamin Khong, IGM 100/42, was previously identified as O. philoceratops ( Barsbold, 1981; Barsbold et al., 1990, fig. 10.1A and B View Fig ), but it is more similar to Citipati in the shape of its premaxilla and circumnarial region and differs from O. philoceratops in the length of the maxilla and dentary (see below). It may represent a second species of Citipati .
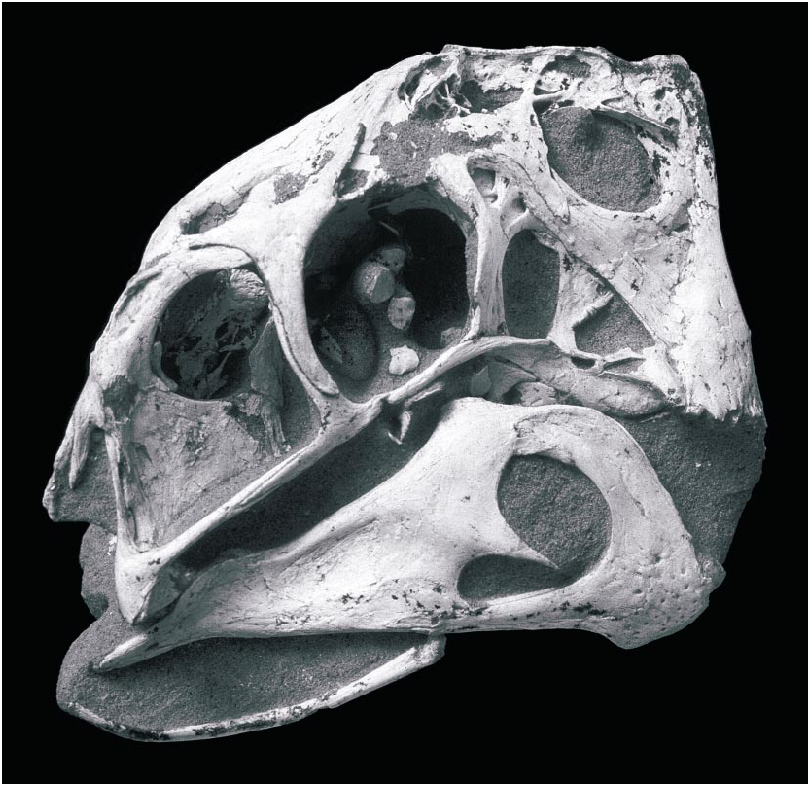
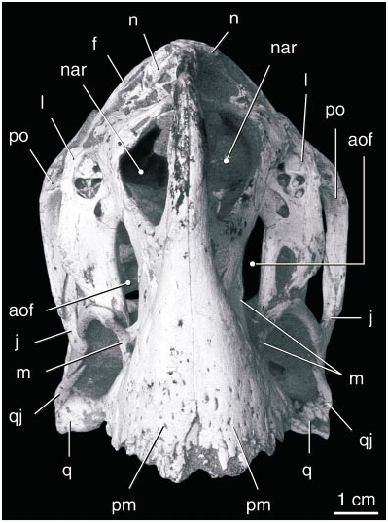
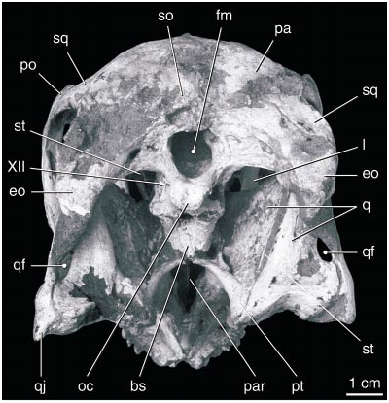
DESCRIPTION: The skull of the holotype ( figs. 1–10 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig ) is complete, including both stapes and the paired elements of the hyoid apparatus. In lateral view ( fig. 2 View Fig ), the skull is almost rectangular, with a vertical premaxilla anteriorly. The right and left premaxillae are fused and edentulous. Ventrally ( fig. 3 View Fig ), the premaxilla expands transversely to form a curved, Ushaped triturating surface in ventral view that supports a series of parallel ridges and troughs. The oral margin of the premaxilla forms a sharp edge that bears a series of large bony denticles, five on each side. Posterior to these, the palatal surface of the premaxilla supports a pair of robust, longitudinal, parasagittal ridges that extend posteriorly along either side of the midline to the posterior edge of the palate. The palatal surface broadens toward the back of the mouth, where it bears two additional ridges that lie lateral and parallel to the first pair.
The large, elliptical nares are bordered ventrally, anteriorly, and anterodorsally by the premaxilla and posteriorly and posterodorsally by the nasal. Owing to profound foreshortening of the face, the naris lies almost entirely dorsal to the antorbital fossa, its rear margin lying nearly as far back as the posterior edge of the antorbital fenestra. The posteriorly fused nasals form a complex array of pockets along the posterodorsal edge of the narial opening. The pockets lie dorsal to a horizontal roof to the nares, and another, narrower horizontal roof overlies these pockets anterodorsally. A broad elliptical opening in the horizontal narial roof communicates between the nares and the dorsal nasal pockets. Dorsal to the naris the nasal forms a vertical lamina, and a lip along its dorsal edge was presumably for the attachment of circumnarial soft tissues.
The large premaxilla forms much of the anterodorsal margin of the antorbital fossa. A fossa on the lateral surface of the ascending ramus anterior to the naris ( fig. 4 View Fig ) is similar to an accessory opening of IGM 100/42 described by Barsbold et al. (1990). Posterior to the naris, the premaxilla extends dorsal to the maxilla to meet the nasal, separating the maxilla from the narial opening. The antorbital fossa is triangular with a vertical posterior edge and horizontal ventral edge. The tall, oval antorbital fenestra is separated from the large, triangular accessory antorbital fenestra by an hourglassshaped, inset interfenestral bar formed by the maxilla. Dorsal to the accessory fenestra are several smaller fenestrae near the dorsal margin of the fossa. The anteroventral margin of the fossa is composed of the maxilla, and a small splint of maxilla extends posterodorsally above the anterior edge of the fossa. Posteriorly, ventral to the antorbital fenestra, the fossa is bordered by the slender anterior process of the jugal.
The lacrimal is transversely broad and forms a concave, posteriorly facing surface along the anterior edge of the orbit. It is perforated by a large lacrimal foramen. The lacrimal canal passed through this foramen, traversing a laterally open trough above the internal antorbital fenestra ( Witmer, 1997) before penetrating the side of the face to enter the nasopharyngeal cavity just behind the posterodorsal extremity of the naris. The anterior part of the lacrimal dorsal to the antorbital fossa houses a broad pneumatic pocket opening anteriorly, within which lies several recessed pneumatic pockets.
There is no evidence of a prefrontal. The paired frontals contact the nasals in a simple suture ( fig. 5 View Fig ). The joint between the frontals is short but highly complex. Posteriorly, the fused parietals divide the frontals for over onehalf of their length. The frontal participates in the supratemporal fenestra, but it lacks the mandibular adductor fossa that deeply excavates the dorsal surface of the frontal in many other nonavian theropods. Laterally, it is overlapped by an anteromedial process of the postorbital. The frontals contact one another along the midline to form the dorsal surface of the anterior end of the braincase. The ventral surface of the frontal forming the dorsal edge of the orbit houses several small pockets that may have been pneumatic.
The orbit is bounded ventrally by the jugal and posteriorly by the postorbital, which meets the ascending process of the jugal to form a postorbital bar. The infratemporal fenestra is subrectangular, with straight anterior and ventral margins and rounded dorsal and posterior margins. The anterior process of the quadratojugal forms most of the ventral border, and the ascending process forms most of the posterior border of the fenestra. The anterior process is longer than the ascending process. In addition, the quadratojugal possesses a distinct posterior process, as in dromaeosaurids. A large quadrate foramen lies between the quadrate and quadratojugal just dorsal to the posterior angle of the infratemporal fenestra ( fig. 6 View Fig ). The quadrate is massive and tightly sutured to the squamosal, quadratojugal, pterygoid, and braincase. The distal articular surface forms two condyles of approximately equal surface and curvature that are separated by a longitudinal groove. The pterygoid flange of the quadrate is tall and extends posterodorsally to meet the short descending process of the squamosal.
The squamosal is a complex bone that forms the entire dorsal margin of the infratemporal fenestra. Medially, it underlies the parietal, extending nearly to the medial edge of the supratemporal fossa. Anteriorly, it lies medial to the postorbital and ventral to its posterior process. The squamosal curves posterolaterally, and its distal end is forked around the external auditory meatus and downturned. A small posttemporal fenestra may be present on the occipital surface between the squamosal and the paroccipital process as seen on the right side, although this may be an artifact. The articulation between the squamosal and the quadrate and quadratojugal is poorly exposed.
The postorbital is a triradiate bone forming most of the postorbital bar. It overlies the frontal anteriorly and the squamosal posteriorly, and it extends lateral to the jugal on the postorbital bar. The laterosphenoid has a short contact with the postorbital at the anterior end of the supratemporal fenestra. The postorbital forms the posterior border of a fossa dorsal to the orbit extending into the pneumatic recesses of the narial region.
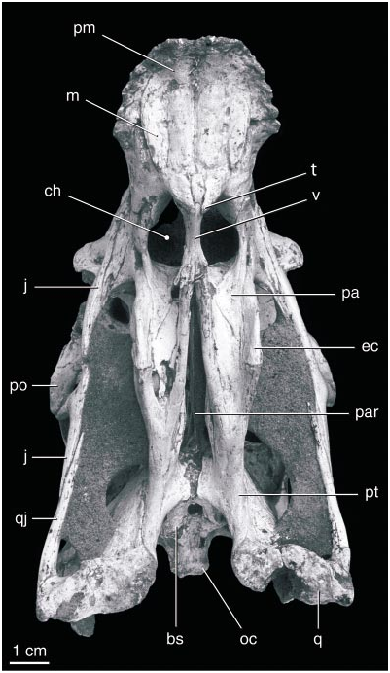
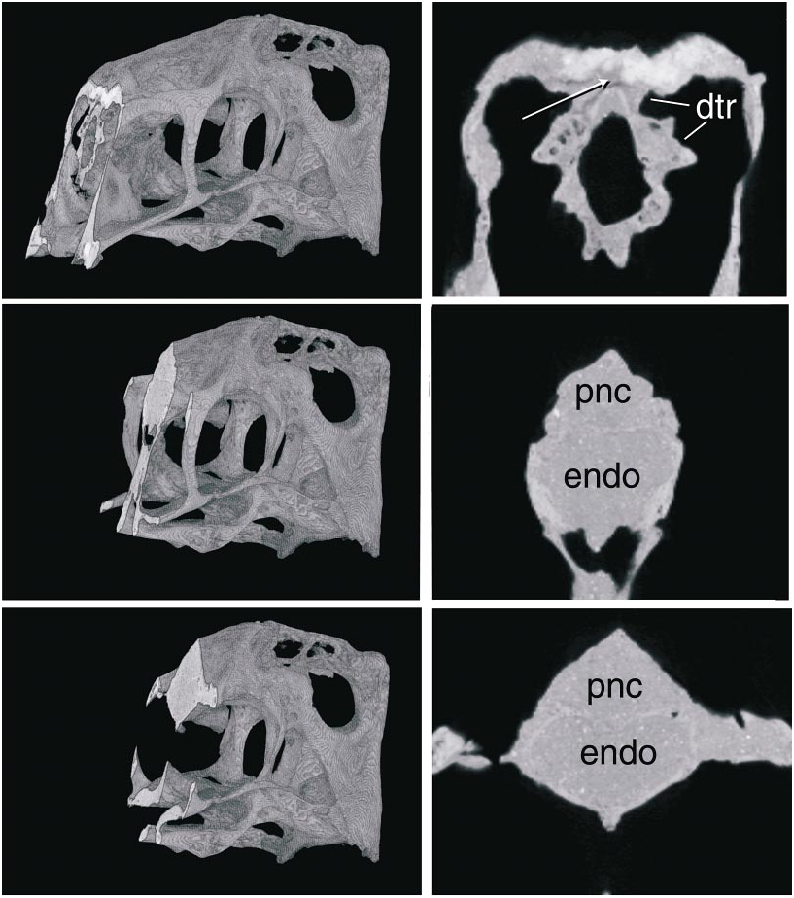
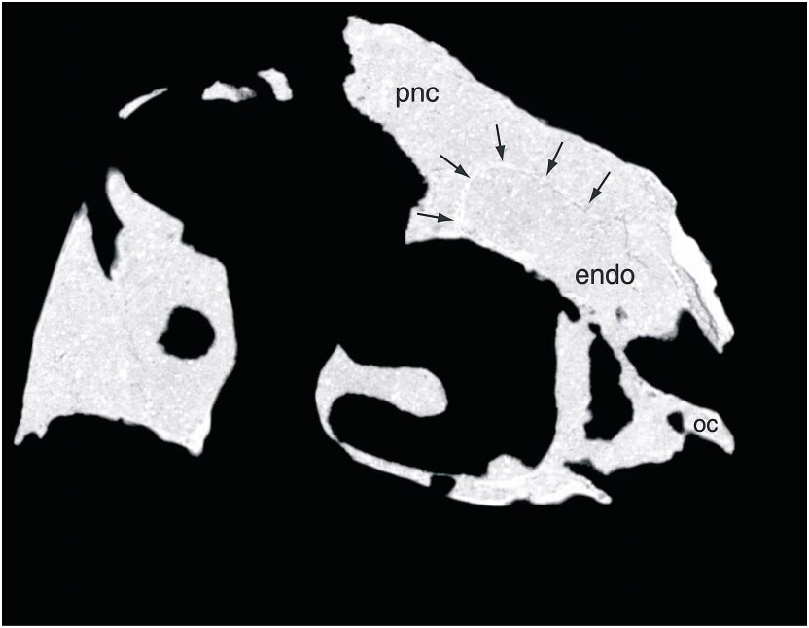
The parietals are fused and form the dorsal surface of nearly half of the skull along the midline ( fig. 5 View Fig ). The fused parietals narrow anteriorly except where they expand slightly at their anterior end. The occipital portion of the parietal is very broad and is oriented obliquely at an angle of nearly 45 ° to face posterodorsally. The parietal forms little of the supratemporal fossa except posteriorly, where a descending flange borders the opening of a dorsal extension of the dorsal tympanic recess. This recess extends anteromedially parallel to the occipital margin immediately posterior to it, and lies almost entirely within the parietal. The CT scans of the skull suggest that the recess connects with a large space within the parietal and frontal dorsal to the brain cavity ( figs. 7 View Fig , 8 View Fig ). The supraoccipital is poorly preserved, but has a low vertical midline crest and may have been separated from the foramen magnum by the exoccipitals.
Within the parietal and frontals lies an extensive space dorsal to the endocranial cavity, as revealed by CT scans ( figs. 7 View Fig , 8 View Fig ). Indeed, this space is larger in volume than the endocranial cavity itself. It is continuous anteriorly with the circumnarial pneumatic cavities and is largest anteriorly, dorsal to the anterior part of the orbit. There is no apparent partitioning of the space, but in the anterior part of the space radioopaque areas laterally indicate what may be internal struts. The space attenuates posteriorly and extends nearly to the occiput ( fig. 8 View Fig ), and it appears to connect with the dorsal tympanic recess above the otic regions of both sides.
The jugal is extremely slender. Posteriorly, it terminates about halfway along the infratemporal fenestra, where it lies lateral to the more robust quadratojugal. It bends anteroventrally anterior to the orbit and extends anteriorly beneath the anterior end of the antorbital fenestra. The right element was damaged beneath the orbit during the life of the animal. The postorbital process of the jugal is robust and forms a broad posterior surface of the orbit. The quadratojugal forms much of the bar beneath the infratemporal fenestra, where it is nearly circular in cross section, and extends anteriorly to beneath the postorbital bar. Dorsally, the quadratojugal extends to meet the squamosal, and it borders the quadrate foramen laterally.
The facial portion of the maxilla is very small and edentulous but includes a single osseous denticle in line with the premaxillary denticles. Posterior to the facial portion, the maxilla forms a narrow shelf ventral to the antorbital fossa. Below this shelf are a variable number of large, anteroposteriorly elongate openings in the ventrolateral surface of the maxilla, two on the right side and one on the left. Posteriorly, the maxilla contacts first the palatine and then the ectopterygoid along its ventral edge. Posterolaterally, the maxilla is overlain by the jugal.
Within the antorbital fossa, the maxilla forms the medial wall. A slender posterodorsal process underlies the premaxilla and forms the dorsal edge of the accessory antorbital fenestra. The nasal overlies the posterodorsal part of the maxilla laterally, and the premaxilla in turn overlies the lacrimal laterally. The interfenestral bar was described above.
The palate ( fig. 3 View Fig ) is generally similar to that of an oviraptorid described by Elzanowski (1999). The lateral portion of the palate is oriented ventrolaterally, and medially the palatal portion forms a distinct posteroventrally directed midline process that surrounds the anterior end of the vomer. The maxilla forms most of the palate anterior to this, which bears four stout longitudinal ridges. A deep groove borders the lateral edge of the lateral ridge. The large choanae lie almost directly beneath the naris and antorbital fossa, such that the nasopharyngeal passageway was oriented almost vertically. The choanae are divided by the vomer posterior to the maxillae.
The palatines are anteroposteriorly short and highly modified. They meet on the midline along the posterior edge of the choanae, where their anterior edge curves dorsally to become vertical and transversely oriented. Laterally, the palatine curves dorsally to meet the maxilla and is separated from the anterior margin of the ectopterygoid by a small, circular, nearly laterally facing suborbital fenestra (the ‘‘postpalatine fenestra’’ of Elzanowski, 1999). The ectopterygoid is vertical, meeting the maxilla laterally and the pterygoid medially and abutting the palatine throughout most of its length. The vomers are fused into a single short, solid bone that does not extend posterior beyond its contact with the pterygoid and palatine at the posterior edge of the choana.
The pterygoids have elongate palatal processes that do not appear to meet along the midline until they contact the vomers. The palatal surface of each element has a longitudinal concavity, confluent anteriorly with the choana. The pterygoid flange is greatly reduced compared to theropods with teeth. It is anteroposteriorly elongate, and, in lateral view, it has a linear dorsal edge and a ventrally convex, crescentic ventral edge, the ectopterygoid forming the anterior half. The flange does not extend laterally beyond the body of the pterygoid, indicating that the M. pterygoideus was relatively small. There is no evidence for a subsidiary palatal fenestra between the pterygoid and palatine. Posteriorly, the pterygoids converge towards the midline at the basipterygoid joint, but they do not come into contact, suggesting that if a contact was present it was formed by soft tissue. The posteromedial edge of the pterygoid is expanded near the midline, as described by Elzanowski (1999). The quadrate ramus of the pterygoid is anteroposteriorly short but very tall, and with the quadrate forms the lateral surface of a deep pocket lateral to the braincase that narrows anteriorly. The pterygoid extends posteriorly to end medial to the mandibular articulating surface of the quadrate. A small, dorsomedially opening pocket is formed dorsal to the posterior end at the level of the quadrate foramen. Another pocket on the dorsal surface of the pterygoid lateral to the basipterygoid joint extends anteriorly into the body of the pterygoid.
An isolated oviraptorid quadrate was described by Maryanska and Osmólska (1997), and the quadrate of Citipati is generally similar. It is oriented obliquely in lateral view, extending posteroventrally from the squamosal. The quadrate has a mediolaterally biconvex mandibular trochlea. The vertical body of the quadrate narrows dorsally, but its dorsal articulation with the squamosal and, apparently, the paroccipital process is not exposed. The pterygoid ramus of the quadrate is very tall, equal in height to about half that of the occipital region.
An epipterygoid is present on both sides of the skull. It is much broader anteroposteriorly than it is thick. It has a broad articulation ventrally with the pterygoid, where it faces anteromedially, and it twists laterally as it narrows dorsally. Dorsally, it contacts the ventrolateral surface of the coossified braincase where a vertical ridge on the laterosphenoid converges with a ridge along the capitate process. On the left side it appears to be fused to the braincase, but not on the right. The dorsal tip twists dorsolaterally and is expanded in the transverse plane, aligned with the vertical ridge on the laterosphenoid above it.
The elements comprising the braincase are coossified, although some areas of contact between bones are apparent. The basioccipital is notable for the posterior extension of the occipital condyle well beyond the posterior end of the foramen magnum ( fig. 5 View Fig ). Indeed, with the floor of the braincase horizontal most if not all of the basioccipital lies posterior to the dorsal edge of the foramen magnum due to the anterodorsal slope of the occiput. The condyle has a nearly flat posterior surface with a central depression. The basioccipital has a midline depression on its ventral surface. The foramen magnum is nearly circular except for a ventral midline depression ( fig. 6 View Fig ). The lateral surface of the basioccipital is concave, and at least three small openings within the concavity presumably enter a pneumatic recess in the body of the bone. A large opening in this region on the right side is presumably due to the breakage of thinwalled bone covering a cavity.
A large vacuity is present on the ventral midline between the basioccipital and basisphenoid. The foramen lies mainly within the basisphenoid, dividing it in two posteriorly. The basisphenoid and basioccipital expand at their contact, and on the right side they appear to be fused. The ventral surface of the basisphenoid descends anteroventrally from this contact. Basipterygoid processes are absent, although a matrixfilled area between the basisphenoid and pterygoids suggests that poorly ossified processes may have been present. The basisphenoid has several pneumatic openings and cavities on the lateral surface of the braincase ventral to the trigeminal foramen. These are poorly exposed, covered laterally by the epipterygoid and beneath matrix ventrally. Several delicate struts span these openings.
A delicate, tall parasphenoid process extends above the length of the interpterygoid vacuity, terminating slightly posterior to the level of the vomer. It descends and tapers anteroventrally from its tall posterior base. The lateral surface of the base has a shallow depression dorsally that opens anterodorsally, presumably the site of origin of an ocular muscle, perhaps the M. rectus oculi ventralis.
The laterosphenoid portion of the braincase forms a ventral floor anteriorly, and a discrete orbitosphenoid ossification is present anterior to it. The large, horizontal orbitosphenoids encircle the anterior part of the endocranial cavity, meeting on the midline dorsally and ventrally. Posterior to the orbitosphenoid the laterosphenoid sends a short, ventrally expanding process ventrally along the midline. This process forms the anterior edge of a large foramen, presumably for cranial nerve (CN) II, that is open ventrally. A posterior descending process forms the posterior edge of this foramen and nearly contacts the top of the parasphenoid rostrum. The capitate process of the laterosphenoid is long and slender and extends laterally along the ventral surface of the frontal and contacts the postorbital. A small slit separates the process from the frontal anteriorly. A sharp ridge descends posteromedially from the capitate process to the epipterygoid contact. A vertical ridge within the supratemporal fossa presumably separated portions of the M. adductor mandibulae . The dorsal contact with the frontal and parietal is apparent but posteriorly the laterosphenoid is fused to the prootic. The posterodorsal contact with the parietal slopes anterodorsally parallel to the occiput. The trigeminal opening (CN V) is relatively small compared with dromaeosaurids (e.g., Velociraptor, IGM 100/976) and is smaller than CN II of this specimen. It is slightly longer than tall and exits ventrolaterally from the braincase. The lateral surface of the laterosphenoid is rugose, presumably indicating an origin of adductor muscles.
The prootic has a welldeveloped fossa dorsally, homologous to the dorsal tympanic recess described in dromaeosaurids ( Norell and Makovicky, 1997, 1999). A horizontal swelling forms the ventrolateral border of the fossa. Dorsal to this fossa lies a large pneumatic opening extending medially into the braincase, as described above. The prootic apparently surrounds this opening dorsally, separating it from the parietal.
The opening in the prootic for CN VII is well preserved on the right side, but on the left it is apparently within a crushed area just ventral to the horizontal swelling. On the right side a depression with at least two, presumably pneumatic, openings lies dorsal to the foramen for CN VII, but this depression is absent on the left side. The otic recess is poorly preserved on both sides, and the fenestrae ovale and rotundum cannot be identified within it. The recess is larger on the right side than the left, and both are small compared with other coelurosaurs.
The contacts between the opisthotic and the prootic and basioccipital appear to be fused. A posterior tympanic recess is present within the anterior surface of the paroccipital process on the right side, but this region is broken on the left. The recess continues medially into the braincase from within a shallow fossa. The fossa penetrates the posterior surface of the paroccipital process via a narrow, mediolaterally elongate opening. In medial view, the jugular foramen and the exits of two hypoglossal foramina are visible within the braincase. The hypoglossal foramina have a short, simple course ventrolaterally from the foramen magnum, opening into a shallow depression. Two ventrolaterally opening foramina on the ventral edge of the paroccipital process presumably originate within the metotic foramen, and likely conveyed CN IX and X and the associated vasculature. The more ventromedial of the two is larger and may be the foramen vagi. The more lateral foramen is small, similar in size to the hypoglossal foramina, and possibly for CN IX. The paroccipital processes are pendant and curve medioventrally, attenuating distally.
Both stapes are preserved, the left element incompletely and the right disarticulated from the skull. The stapes is elongate, straight, and slender, with a footplate that is approximately three times the diameter of the shaft. As implied by the position of the otic notch in the squamosal, the stapes would have projected posterolaterally from the fenestra ovalis at an angle of about 45 ° and slightly ventrally.
Five scleral ossicles, from a partial scleral ring, preserved in the right orbit and below it were removed during preparation ( fig. 1 View Fig ). They are roughly square in shape with irregular margins and a gently concave medial surface. They are similar to those of other basal coelurosaurs such as troodontids and dromaeosaurids.
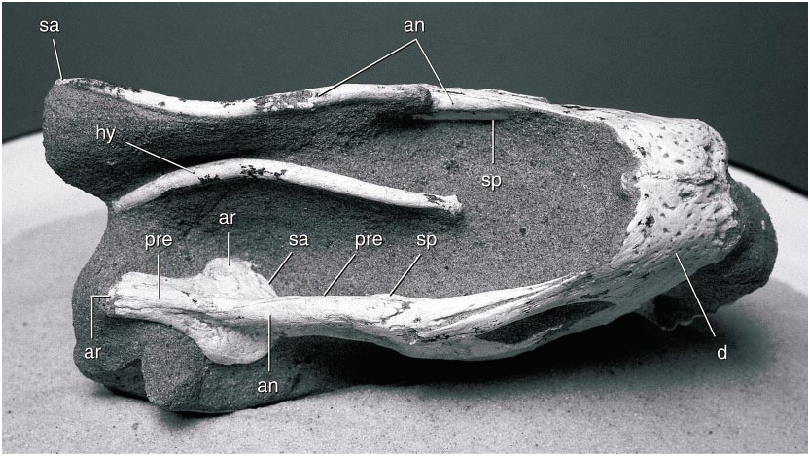
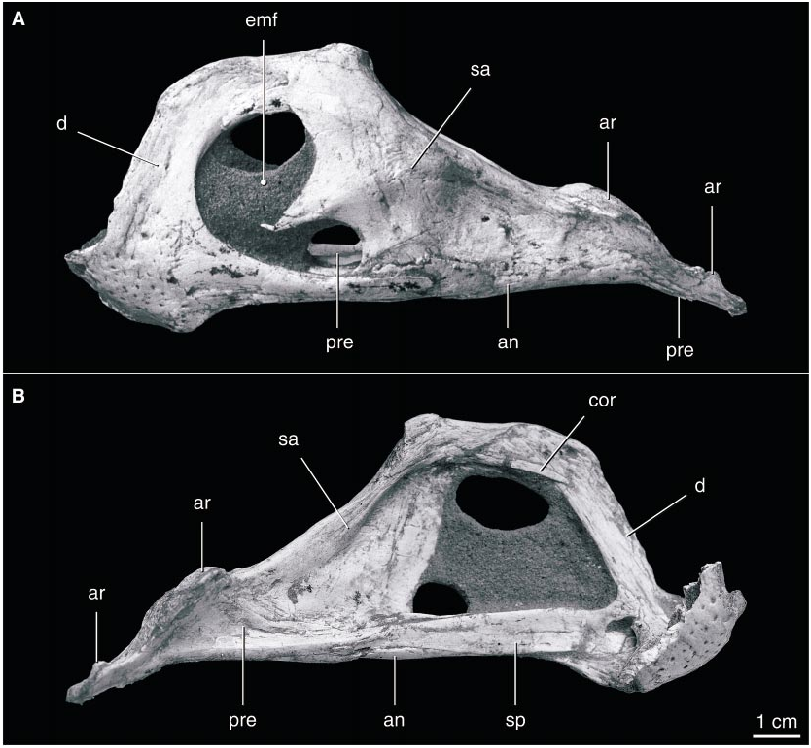
The edentulous mandible ( fig. 9 View Fig ) is dominated by a high, arching coronoid eminence. The symphysis is short and broad, with a transversely oriented, upturned anterior edge. It is slightly below the level of the mandibular articulation. The dorsal surface of the symphysis is concave dorsally behind the anterior edge, and the posterior edge of the symphysis descends posteroventrally. The coronoid eminence rises abruptly from the posterior part of the symphysis and flattens dorsally. The surangular descends gradually from the eminence and extends posteriorly to the end of the mandible, completely covering the lateral surface of the articular. The large mandibular fenestra is divided by an anterior process of the surangular that reaches approximately threequarters of the distance across the fenestra. The lateral surface of the surangular is gently depressed anterior to the mandibular articulation. The adductor fossa on the medial surface of the mandibular ramus is extremely large and bordered ventrally by a slender splenial. The anterior end of the splenial is divided into dorsal and ventral rami in the symphysis. It overlaps the prearticular in the middle of the mandibular ramus, and the prearticular extends posteriorly to the end of the mandible, covering most of the medial surface of the articular.
The mandibular articulation surface is anteroposteriorly elongate and unbounded anteriorly or posteriorly, suggesting that the mandible was capable of anteroposterior movement (cf. Osmólska, 1976). The surface bears a longitudinal midline ridge and faces posterodorsally. The retroarticular process descends posteroventrally from the articular surface, with which it is coplanar, and its ventral edge descends slightly below the level of the remainder of the mandible. The mandibular articulation is much wider than the remainder of the mandibular ramus, and it is bordered ventrally by the prearticular medially and the surangular laterally.
A very small coronoid element appears to be present. A splint of bone on the medial surface of the posterodorsal ramus of the dentary on both sides is in the position of a coronoid bone, although this bone is reported as absent in other oviraptorids ( Barsbold et al., 1990). A bone as small as this could be easily lost or overlooked in poorly preserved specimens, suggesting that it may also be present in other oviraptorids.
A pair of elements of the hyoid apparatus were preserved beneath the mandible ( fig. 10 View Fig ), presumably the ceratohyals (cornua branchiala I). They are simple structures, essentially elongate rods that curve dorsally posteriorly. The anterior and posterior ends are expanded and slightly compressed mediolaterally. They were preserved paralleling the ventral edge of the mandible, possibly their natural position.
Most elements of the postcranial skeleton of C. osmolskae were described by Clark et al. (1999) for the referred specimen IGM 100/979. This specimen preserves only a single vertebra, however, and lacks the ilia and most of the scapulae, pubes, and ischia. It differs from Khaan in that the first metacarpal of the latter is reduced proximally and the bone does not expand, whereas that of Citipati is unreduced and expanded.
The postcranial skeleton of the holotype of C. osmolskae is very well preserved and includes representatives of nearly every element, but it has not yet been fully prepared. The cervical vertebrae are distinctly more elongate than those of Khaan and other oviraptorids where known. The cervical ribs are also more elongate, longer than the corresponding centrum in Citipati but shorter than in Khaan .


| IGM |
Geological Institute, Mongolian Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |