
Monstrillopsis cahuitae Suárez-Morales and Carrillo, 2013
|
publication ID |
https://doi.org/10.1080/00222933.2012.742933 |
|
publication LSID |
lsid:zoobank.org:pub:F3B9C990-9318-4B91-A28B-201C4426C67B |
|
persistent identifier |
https://treatment.plazi.org/id/6378878D-7031-B35D-5D1A-FE78FEBBAAEE |
|
treatment provided by |
Felipe |
|
scientific name |
Monstrillopsis cahuitae Suárez-Morales and Carrillo |
| status |
sp. nov. |
Monstrillopsis cahuitae Suárez-Morales and Carrillo , sp. nov.
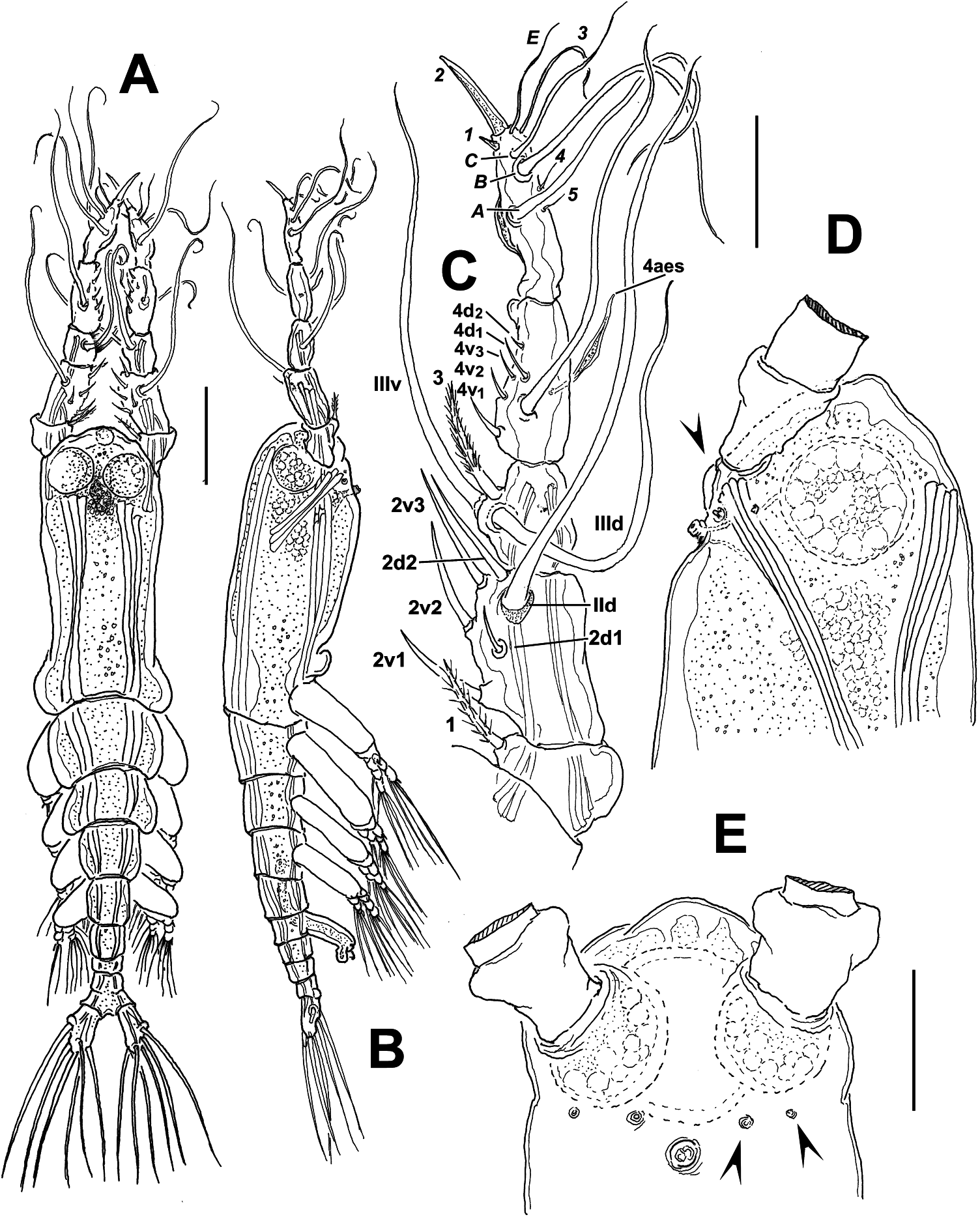
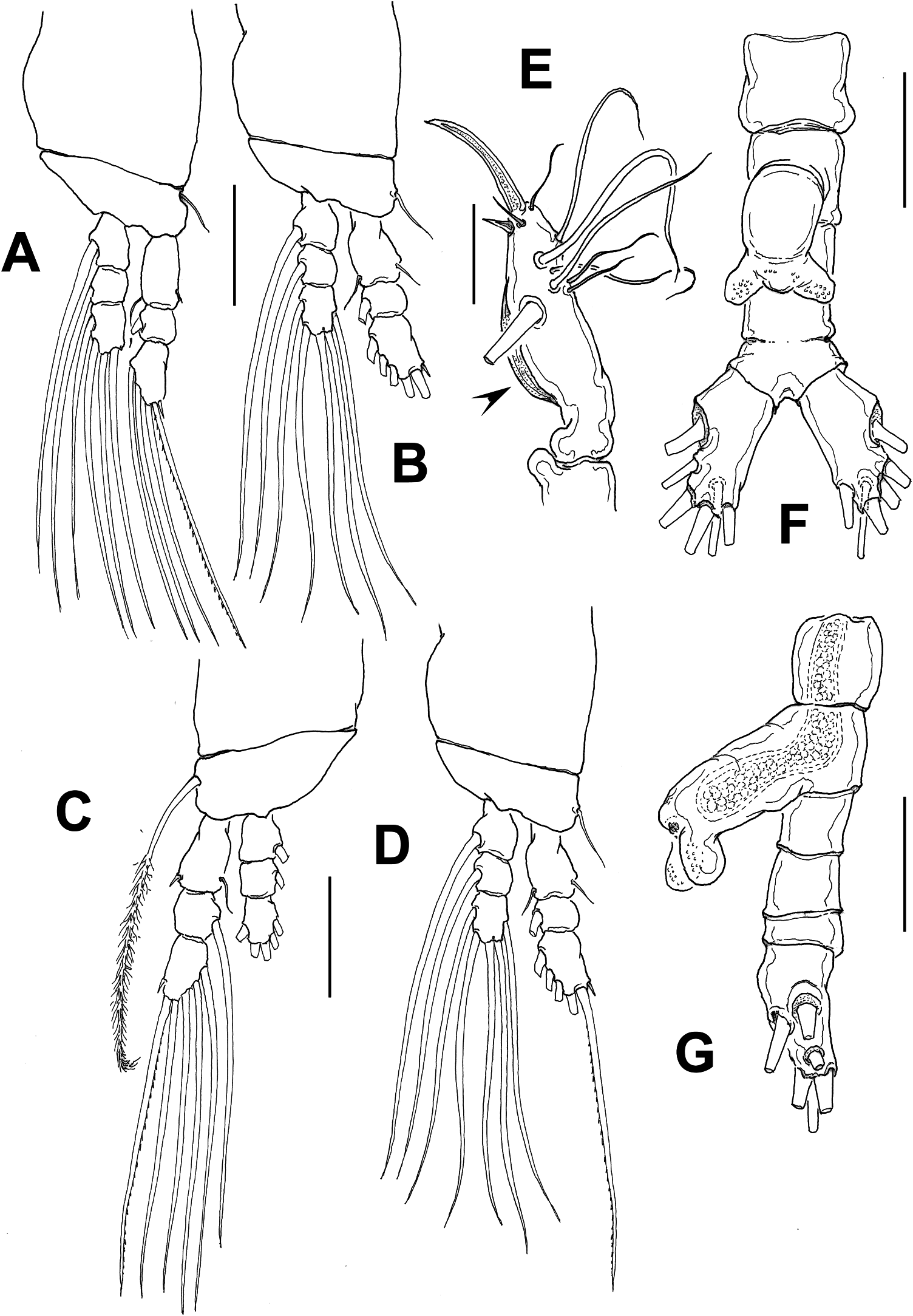
( Figures 4 View Figure 4 , 5 View Figure 5 )
Diagnosis
Small species (0.55 mm), with oral papilla at 20% of way back along cephalothorax, terminal spiniform process less than half the length of bearing segment. Genital apparatus cylindrical, with paired globose processes distally. Six caudal setae.
Material examined
Holotype. Adult male from Cahuita National Park (sta. 2: 9 ◦ 44.31 ′ N, 82 ◦ 48.13 ′ W), Limón, Costa Rica, Central America , partially dissected, ethanol-preserved, vial. Selected appendages on slides mounted on glycerine, sealed with Entellan ®. Date and hour of collection: 11 February 2011, 07:40 h. Plankton sample; depth to bottom: 2 m. Vial and slides deposited in the Collection of Zooplankton at El Colegio de la Frontera Sur ( ECOSUR), in Chetumal, Mexico (ECO-CHZ-07570). GoogleMaps
Description
Total body length of adult male holotype: 0.55 mm, measured from anterior end of cephalothorax to posterior end of anal somite. Cephalothorax 0.26 mm long, representing up to 48.6% of total body length. Antennule 0.27 mm long, representing 92% of the cephalothorax length ( Figure 4A,B View Figure 4 ). Oral papilla reduced, located anteriorly, about 20% of way back along ventral surface of cephalothorax ( Figure 4D View Figure 4 ). Pair of relatively large pigment cups present, moderately developed, separated by less than half length of eye diameter, weakly pigmented; ventral cup larger than lateral cups. Forehead with moderate medial rounded protrusion. Wide-based, low rounded process on ventral surface protruding between bases of antennules (arrowed in Figure 4D View Figure 4 ). Two pairs of nipple-like cuticular processes on anterior ventral surface between antennule bases and oral papilla (arrowed in Figure 4E View Figure 4 ).
As usual in male monstrilloids, antennules five-segmented, geniculate ( Figure 4C View Figure 4 ). In terms of pattern described by Grygier and Ohtsuka (1995), element 1 present on first segment, biserially setulated; elements 2d 1, 2d 2, 2v 1, 2v 2, 2v 3 and IId on second segment. Third segment with elements 3, IIId and IIIv. Segment four bearing elements 4d 1,2 and 4v 1–3 as well as IVd. Aesthetasc 4aes on ventral surface. Following Huys et al. (2007) nomenclature for male monstrilloids’ terminal segment, armature as: elements 1–7 present on anterior margin, unbranched elements A and B on posterior margin. As usual in male Monstrillopsis , terminal antennular segment modified, with inner rounded expansion and elongate distal half, forming a sabre-like structure, curved distally ( Figures 4C View Figure 4 , 5E View Figure 5 ).
First pedigerous somite incorporated into cephalothorax; this and succeeding three free pedigerous somites each bearing pair of biramous swimming legs. Pedigerous somites 2–4 together accounting for 38% of total body length in dorsal view. Intercoxal sclerites of legs 1–4 sub-rectangular, without ornamentation on surface or along distal margin. Basis of legs articulating with rectangular coxa along diagonal line. Basis with thin lateral seta on legs 1–4; on leg 3, this seta thicker and 4.5–5 times longer than in other legs, reaching well beyond distal margin of exopodal ramus ( Figure 5C View Figure 5 ); heavily setulated from distal two-thirds. Endopodites and exopodites of swimming legs 1–4 triarticulated ( Figure 5A–D View Figure 5 ). Ramus setae all lightly and biserially plumose except spiniform outer seta on exopodal segments 1 and 3, and inner seta of first exopodal segment, these latter being short, slender. Also, outer apical exopodal seta of swimming legs 1–4 with outer margin smooth, inner margin lightly spinulose.
Armature formula of swimming legs is shown in Table 2.
Fifth legs absent. Urosome consisting of five somites: fifth pedigerous somite, genital somite with genital apparatus, two free postgenital somites, and partially fused (dorsal segmentation division clear, weaker on ventral view) anal somite. Ventral surface of genital somite forming enlarged base of cylindrical shaft with distal genital lappets. Lappets represented by pair of bulbous processes posteriorly directed, moderately divergent ( Figure 5F,G View Figure 5 ), distal surface of lappets with clusters of blisterlike cuticular processes. Caudal rami subrectangular, weakly divergent, approximately 1.6 times longer than wide, each ramus bearing six setae. Inner dorsal seta (seta VII in Huys and Boxshall 1991) thinnest.
Type locality
Cahuita National Park , Caribbean Coast of Costa Rica (9 ◦ 45 ′ N, 82 ◦ 49 ′ W) GoogleMaps .
Etymology
The specific epithet makes reference to the National Park of Cahuita, Costa Rica, the type locality of this species .
Remarks
The male specimen collected from Cahuita, Costa Rica is assignable to the genus Monstrillopsis by its possession of a modified fifth antennular segment, with an inner rounded protuberance and an attenuated, sabre-like distal half; this kind of modification occurs only in males of Monstrillopsis (see Huys and Boxshall 1991; Suárez-Morales et al. 2006). Also, the position of the oral papilla near the anteriormost part of the cephalothorax and fully developed eyes are present in this specimen; both characters are regarded as diagnostic of the genus ( Sars 1921; Suárez-Morales et al. 2006). The presence of six caudal setae in our specimen of Monstrillopsis is quite unexpected. This is usually regarded as a character of the genus Monstrilla and having four such setae is associated with Monstrillopsis ( Sars 1921; Suárez-Morales and Ivanenko 2004; Suárez-Morales et al. 2006). In this case it was not considered definitive because the number of caudal setae appears to be more consistent in females than in males; for instance, both the female and the male of Monstrillopsis zernowi bear five caudal setae ( Dolgopolskaya 1948) and represent “aberrant” cases among the members of this genus ( Suárez-Morales et al. 2006). The decision to assign this male to Monstrillopsis is supported by the strongest character for males of this genus: the unique morphology of the last antennular segment, as described by Huys and Boxshall (1991).
Of course, this unique combination of characters is not present in any other known congener of Monstrillopsis and so the specimen from Costa Rica is recognized as a new species. Another striking character of this species is its small size (body: 0. 55 mm long); it is among the smallest monstrilloids known. Only Monstrilla pygmaea Suárez-Morales, 2000 (0.43 mm) and Monstrilla minuta Isaac, 1975 (0.49 mm) are smaller. Besides the antennular modification another interesting character shared by males of Monstrilla and Monstrillopsis is the presence of a long, cylindrical genital apparatus with paired terminal globose processes. This kind of structure has been recorded mainly in species of Monstrilla (i.e. Monstrilla reidae Suárez-Morales, 1993 and Monstrilla globosa Suárez-Morales, 2003 from the Caribbean, Monstrilla bahiana Suárez-Morales and Dias, 2001 from Brazil, Monstrilla patagonica Suárez- Morales, 2008 from the Beagle Channel). It is not exclusive of the genus because it has also been described in species of Monstrillopsis , as in Monstrillopsis zernowi Dolgopolskaya, 1948 from the Black Sea, Monstrillopsis fosshageni Suárez-Morales and Dias, 2001 from Brazil, and in Monstrillopsis chathamensis Suárez-Morales and Morales-Ramírez, 2009 from the Pacific coast of Costa Rica ( Suárez-Morales and Dias 2001; Suárez-Morales and Morales-Ramírez 2009; see also Huys and Boxshall 1991, figs. 2.5.8C,D). A different kind of genital apparatus reported in species of Monstrillopsis is that described in Monstrillopsis chilensis Suárez-Morales, Bello-Smith and Palma, 2006 from the Beagle Channel, with long genital lappets (Suárez-Morales et al. 2008).
The Costa Rican specimen is comparable to Monstrillopsis fosshageni , but differs in being much smaller (0.55 mm versus 1.8 mm), having a terminal spiniform process of the fifth antennular segment less than half the length of the segment versus a longer element in Monstrillopsis fosshageni , representing more than half the length of the segment. Although some antennular elements are absent in Monstrillopsis fosshageni (see Suárez-Morales and Dias 2001), in this species element 1 is extremely long and heavily setulated whereas it is relatively short and lightly setulated in the new species. The genital apparatus has the same structure and proportions, but the terminal lappets are relatively stronger in the new species; also, in Monstrillopsis fosshageni , these are smooth, without the blister-like ornamentations described in Monstrillopsis cahuitae sp. nov.
Suárez-Morales et al. (2006) included the following nominal species in the genus Monstrillopsis : Monstrillopsis dubia ( Scott, 1904) from Scotland (60 ◦ N), Monstrillopsis zernowi from the Black Sea (43 ◦ N), Monstrillopsis sarsi Isaac, 1974 from England (54 ◦ N), Monstrillopsis fosshageni from Brazil (20 ◦ S), Monstrillopsis dubioides Suárez-Morales, 2004 from Norway (62 ◦ N), Monstrillopsis ferrarii Suárez-Morales and Ivanenko from the White Sea in the Arctic Ocean (66 ◦ N), Monstrillopsis chilensis from off Chile (33 ◦ S), Monstrillopsis igniterra from the Southern Ocean (55 ◦ S). Later on, Suárez-Morales and Morales-Ramírez added Monstrillopsis chathamensis from the Eastern Pacific. Together with Monstrillopsis fosshageni , the new species Monstrillopsis cahuitae is the second Monstrillopsis species known from a male only. Only four of the ten species of Monstrillopsis has been recorded from subtropical or tropical areas. Hence, it is expected that the description of this species based on the male will help the identification of the female in the same reef area.
| ECOSUR |
El Colegio de la Frontera Sur (Mexico) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |