
Gracixalus lumarius, Rowley & Le & Dau & Hoang & Cao, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3785.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:E515AAC3-811B-48BA-A680-E4851887633F |
|
DOI |
https://doi.org/10.5281/zenodo.5062526 |
|
persistent identifier |
https://treatment.plazi.org/id/636E878B-FF99-AB78-41C5-3011FEFFFDE7 |
|
treatment provided by |
Felipe |
|
scientific name |
Gracixalus lumarius |
| status |
sp. nov. |
Gracixalus lumarius View in CoL sp. nov.
Holotype: AMS R 176202 , adult male, in tree hole (approx. 6 cm diameter entrance, 20 cm deep) with eggs, in montane evergreen forest in Ngoc Linh Nature Reserve , Dak Glei District, Kon Tum Province, Vietnam (15.063º N, 107.786º E, 1845 m; Fig. 1 View FIGURE 1 ). Collected at 20:55 h on 3 April 2010 by J.J.L. Rowley, D. T. T. Le, V. Q. Dau and H.D. Hoang. GoogleMaps
Paratypes: UNS 00341 / AMS R 173838 , female, collected on a tree branch approximately 2 m above ground in montane evergreen forest in Ngoc Linh Nature Reserve , Dak Glei District, Kon Tum Province, Vietnam (15.0785º N, 107.971º E, 2160 m; Fig. 1 View FIGURE 1 ), collected at 21:30 h on 16 July 2009 by D GoogleMaps . T. T. Le and V.Q. Dau . AMS R 173889 and AMS R 173890 , adult males collected on tree branch in close proximity, in montane evergreen forest in Ngoc Linh Nature Reserve , Dak Glei District, Kon Tum Province, Vietnam (15.077958º N, 107.960757º E, 1990 m; Fig. 1 View FIGURE 1 ), collected at night on 17 July 2009 by D GoogleMaps . T. T. Le and V.Q. Dau . UNS 00340 / AMS R 176203 , juvenile, on plant leaf 1 m above ground in montane evergreen/bamboo forest in Ngoc Linh Nature Reserve , Dak Glei District, Kon Tum Province, Vietnam (15.063º N, 107.862º E, 2035 m; Fig. 1 View FIGURE 1 ), collected at 20:35 h on 3 April 2010 by J.J.L. Rowley, D GoogleMaps . T. T. Le , V.Q. Dau and H.D. Hoang .
Other material. Eggs collected from same tree hole and at the same time and date as the holotype, AMS R 176213 ( Fig. 7B View FIGURE 7 ) .
Etymology. Specific name from lumarius L., meaning of thorns, in reference to the extensive conical tubercles over the dorsal surface of the new species.
Suggested common name. Thorny tree frog (English), Ếch cây gai (Vietnamese).
Diagnosis. The new species is assigned to the family Rhacophoridae by the presence of intercalary cartilage between the terminal and penultimate phalanges of digits, tips of digits expanded into large disks bearing circummarginal grooves, vomerine teeth absent and horizontal pupil ( Liem 1970; Duellman & Trueb 1986; Brown & Alcala 1994). Although morphological characters for the genus require revision, the species is allocated to the genus Gracixalus by the presence of spines on the upper eyelid, distance between nostrils less than between eyes, rictal gland connected to the mouth, tibia length greater than four times width ( Delorme et al. 2005), back bearing dark X-or inverted V-shape ( Fei et al. 2009) and by molecular data (see below). Gracixalus lumarius sp. nov. is distinguished from other rhacophorids in Indochina by a combination of (1) medium body size (adult males 38.9– 41.6 mm; adult female 36.3 mm), (2) dorsum brown diurnally and yellow nocturnally, (3) venter pink, (4) tympanum and supratympanic fold indistinct (5) iris dark gold with a dense, relatively uniformly distributed network of black reticulations, (6) dorsum with distinctive white conical tubercles in adult males, and (7) eggs deposited on wall of a phyptotelm.
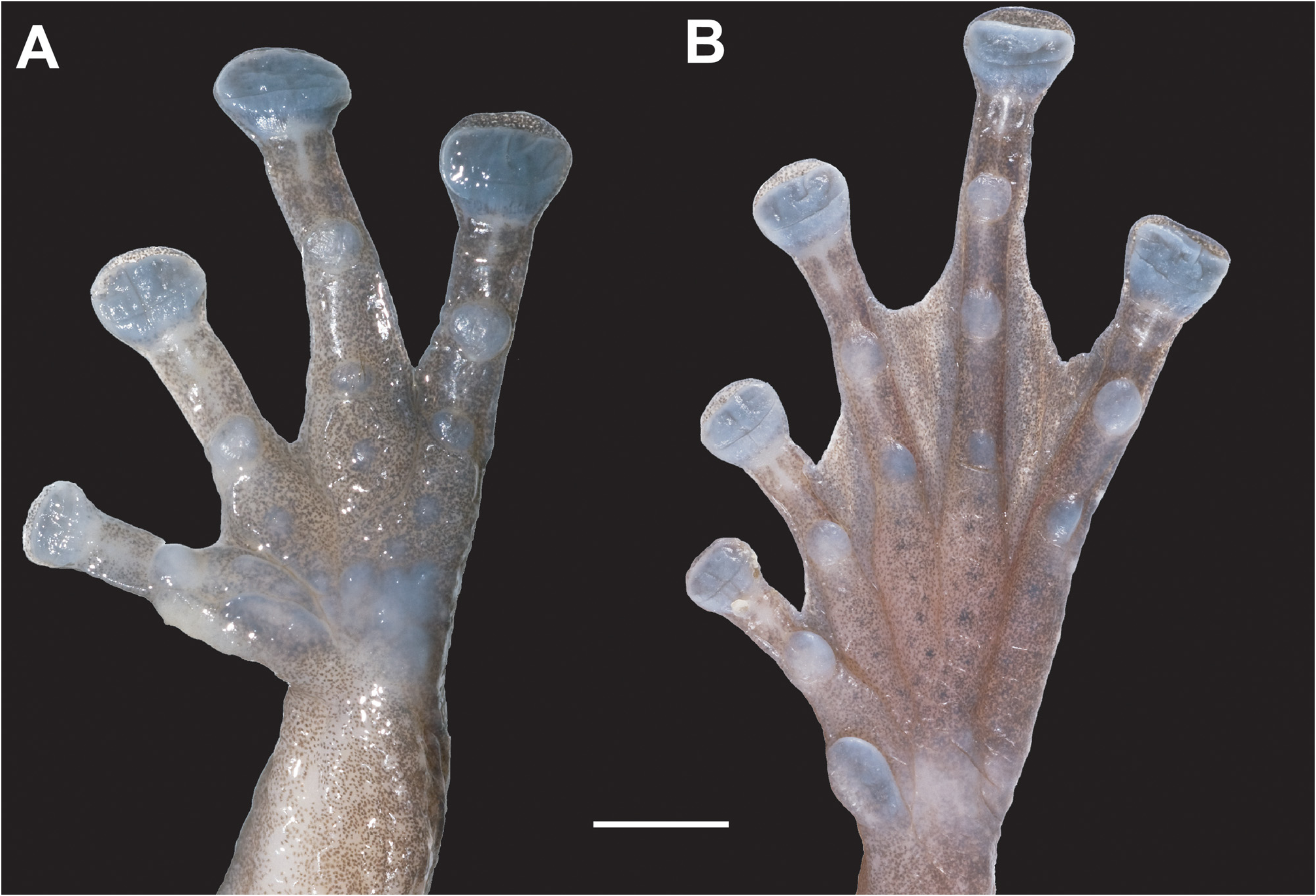
Description of holotype. Medium-sized rhacophorid (41.6 mm), body robust, head length 89% of head width, snout rounded in dorsal view, gently truncate in profile, projecting slightly beyond margin of lower jaw, canthus rostralis indistinct, rounded, loreal region straight, interorbital region convex, nostrils oval, slightly protuberant, without flap of skin laterally, slightly closer to tip of snout than eye, pupil oval, horizontal, tympanum indistinct, approximately one third of eye diameter, pineal ocellus absent, skin not co-ossified to forehead, vomerine teeth absent, choanae oval, at margins of roof of mouth, tongue attached anteriorly, deeply notched posteriorly, tooth-like projections on lower jaw absent, supratympanic fold indistinct. Weakly visible rictal gland connected to the mouth. Vocal sac present, pair of distinct, oval openings at base of jaw, baggy gular region. Forelimbs relatively robust, relative length of fingers I <II <IV <III. Tips of all fingers with well-developed disks with circummarginal grooves; disks relatively wide compared to finger width (third finger disk 204% third finger width), third finger disk width greater (144%) than tympanum diameter; slight dermal fringes on fingers, basal webbing at base of fingers II–IV. Subarticular tubercles prominent, rounded, formula 1, 1, 2, 2. Accessory palmar tubercles distinct; palmar tubercle irregular, flat; thenar tubercle indistinct; prepollex elongate, with low, oval tubercle; distinct, nuptial pads present on inner surface of the prepollex ( Fig. 4A View FIGURE 4 ). Relative length of toes I <II <V <III <IV, tips of toes with well-developed disks with distinct circummarginal grooves, wide when compared to toe width. Webbing present, formula I 1 ½ – 2 II 1 ⅓ – 2 + III 1 ½ – 2 + IV 2 + – 1 + V, with dermal fringes, subarticular tubercles prominent, rounded, formula 1, 1, 2, 3, 2, inner metatarsal tubercle distinct, outer metatarsal tubercle and supernumary tubercles absent ( Fig. 4B View FIGURE 4 ). Dorsal surface of head and back, including snout, eyelids and tympanum covered in dense network of conical tubercles; except for several tubercles scattered on the dorsal surface of arms, the dorsal surface of forelimbs and hindlimbs are smooth. Ventral surface of thighs and abdomen coarsely granular, throat finely granular. No dermal fringes on limbs, no tibiotarsal projection, supracloacal glands.
Colour of holotype in life. Dorsal surface yellowish brown with a darker brown interorbital crossbar and cross-mark on the back starting between the eye; limbs with faint darker barring. Tips of toes and webbing pinkish grey. Loreal and tympanic markings absent. Flanks bright yellow with pink mottling dorsally, changing to pink with yellow mottling ventrally. Throat, chest, belly and ventral surfaces of limbs pink, slightly paler pink on chest. Small glands around vent creamy whitish. Iris dark gold with a dense, relatively uniformly distributed network of fine black reticulations; iris periphery black; eye periphery grey posteriorly. Dorsal colouration varied in brightness over time.
Colour of holotype in preservative. Dorsal surface as in life, but conical tubercles paler, white, and more distinct. Bright yellow and pink colours faded. Chest and belly pinkish white, margins of throat, ventral surfaces of limbs, hands, feet, including webbing, pale brown.
Measurements. Holotype: SVL 41.6, HDL 14.3, HDW 16.1, SNT 5.4, EYE 5.8, IOD 4.3, TMP 2.0, TEY 0.8, IN 3.9, NS 2.5, EN 2.6, TIB 19.6. ML 14.8, PL 18.8, IMT 2.1.
Variation. Paratypes do not vary considerably in their colouration. In preservative, AMS R 176202 is slightly darker than other specimens. AMS R 173890 has higher contrast patterning. All adult males have a dorsum with distinctive, white conical asperities, but these vary in extent among individuals. Compared to the holotype, AMS R 173889–1738890 have smaller, less raised tubercles covering anterior two-thirds of dorsum, and not extending on to dorsal surface of arms. The single female ( UNS 00341/ AMS R 173838 ) and single juvenile ( UNS 00340/ AMS R 176203 ) specimens have less obvious raised tubercles in place of asperites ( Fig. 5 View FIGURE 5 ). The single juvenile ( UNS 00340/ AMS R 176203 ) is uniformly pale tan without dorsal markings or barring on limbs, but with a slightly darker canthal stripe. Male paratypes lack nuptial pads. Measurements of the type series are shown in Table 2 View TABLE 2 .
* holotype
Eggs. Approximately 130 eggs were deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm above the water (AMS R 176213; Fig. 7B View FIGURE 7 ). The eggs are approximately Gosner (1960) stage 12, contain two jelly layers and measure an average of 2.75 mm (n=3) in diameter in preservative. Ova, which measure ~ 1.3 mm diameter, were coloured bluish grey in life and creamy grey in preservative. The eggs were identical in the mtDNA fragment of sequenced adults.
Ecology. The holotype was found underwater in a tree hole with conspecific eggs on the wall of the phyptotelm ( Fig 7B View FIGURE 7 ). All other specimens were found on arboreal vegetation, away from streams or ponds in montane evergreen or bamboo forest (eg. Fig 7A View FIGURE 7 ). The advertisement call and larvae of the new species are unknown. At present, the species is only known from montane bamboo and evergreen forest (> 1800 m) on Mount Ngoc Linh and adjacent peaks, with less than 20 km between the farthest localities. The distribution of the species is unknown, but is likely to be restricted to high-elevation forest on Mount Ngoc Linh and adjacent peaks.
Comparisons. Gracixalus lumarius sp. nov. is distinguished from all other species of Gracixalus , and all other medium-sized (35–45 mm adult SVL) rhacophorids with brownish dorsum in Indochina by a combination of the following characters: dorsum brown diurnally and yellowish brown nocturnally; venter pink; tympanum and supratympanic fold indistinct; vomerine teeth absent; iris dark gold with a dense, relatively uniformly distributed network of black reticulations; dorsum with distinctive, white conical asperities in males; and eggs deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm.
Within the genus Gracixalus , Gracixalus lumarius sp. nov. is distinguished from Clade I of Rowley et al. (2011), G. gracilipes (Bourret) , G. quangi Rowley, Dau, Nguyen , Cao & Nguyen, G. quyeti (Nguyen, Hendrix, Böhme, Vu & Ziegler) , and G. supercornutus (Orlov, Ho, & Nguyen) by having a dorsum brown diurnally and yellow nocturnally and a pink venter (versus greenish dorsum and white or yellow venter), an indistinct (versus distinct) tympanum and supratympanic fold, a dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright copper or olive-gold iris with limited, non-uniform black reticulations), distinctive white, conical asperities on the dorsum (versus dorsum mostly smooth or slightly tuberculate) in males, and eggs deposited on wall of a phyptotelm (versus on vegetation above pools for G. gracilipes , G. quangi , and G. supercornutus, Rowley pers. obs.; unknown in G. quyeti ). G. lumarius sp. nov. also has a larger body size (38.9– 41.6 mm in three adult males) compared to G. gracilipes , G. quangi , and G. supercornutus (all with male SVL <25 mm).
Gracixalus lumarius sp. nov. is distinguished from G. carinensis (Boulenger) by having a pink (versus white) venter, an indistinct (versus distinct) tympanum and supratympanic fold, dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold iris with limited, non-uniform black reticulations), dorsum with distinctive, white conical asperities (versus dorsum smooth or granulated with small warts on head and anterior part of back only) in males. From G. jinxiuensis (Hu) , the new species is distinguished by having a larger body size (38.9–41.6 mm in three adult males versus 23.5 mm in adult male holotype), indistinct (versus distinct) tympanum and supratympanic fold, dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold iris with limited, non-uniform black reticulations), and distinctive, white conical asperities (versus low, scattered tubercles on dorsum) in males. From G. medogensis (Ye & Hu) , the new species is distinguished by having a larger body size (38.9–41.6 mm in three adult males compared to 26.5 mm in one adult male), pink (versus light grey or whitish) venter, indistinct (versus distinct) tympanum and supratympanic fold, and distinctive, white conical asperities (versus smooth) in males. From the recently described G. nonggangensis Mo, Zhang, Luo & Chen and G. waza Nguyen, Le, Pham, Nguyen, Bonkowski & Ziegler , the new species differs by having a larger body size (38.9–41.6 mm in three adult males versus <36 mm in adult males), a pink venter (versus white venter with brown spots), indistinct (versus distinct) tympanum and supratympanic fold, dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus olive iris with limited, non-uniform brown reticulations), and dorsum with distinctive, white conical asperities (versus dorsum smooth or with some small tubercles) in males.
The new species differs from Kurixalus ananjevae (Matsui & Orlov) , which is morphologically and likely molecularly similar to Gracixalus Clade II ( Rowley et al. 2011), by having a larger body size (38.9–41.6 mm in three adult males compared to 32.4 mm in one adult male), pink (versus greyish white) venter, indistinct (versus distinct) tympanum and supratympanic fold, and dorsum with distinctive, white conical asperities (versus dorsum with irregularly distributed, small tubercles) in males.
Gracixalus lumarius sp. nov. also differs from all other medium-sized (35–45 mm adult SVL) rhacophorids from Indochina. From Kurixalus banaensis (Bourret) , the new species is distinguished by having a pink (versus white) venter, indistinct (versus distinct) tympanum and supratympanic fold, dorsum with distinctive, white conical asperities (versus dorsum granular) in males, no dermal fringing along limbs (versus marked fringing along forearm and tarsus). From Kurixalus odontotarsus (Ye & Fei) , the new species differs by having a pink venter (versus white venter with black or grey marbling), indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), and dorsum with distinctive, white conical asperities (versus dorsum rugose) in males. From Polypedates leucomystax (Gravenhorst) and P. megacephalus Hallowell , the new species differs by having a pink (versus white) venter, indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold iris lacking distinct black reticulations), dorsum with distinctive, white conical asperities (versus dorsum smooth) in males and eggs deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm (versus eggs in foam nest). From Rhacophorus bipunctatus Ahl the new species differs by having a pink (versus yellow) venter, indistinct (versus distinct) supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold iris with limited, non-uniform black reticulations), dorsum with distinctive, white conical asperities (versus dorsum smooth) in males and eggs deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm (versus eggs in foam nest). From R. calcaneus Smith , the new species differs by having a brown dorsum diurnally and yellow nocturnally (versus green or deep reddish-brown dorsum), pink (versus yellow) venter, indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus iris with a bright red periphery), and dorsum with distinctive, white conical asperities (versus dorsum smooth) in males. From R. orlovi Ziegler & Köhler , the new species differs by having a brown dorsum diurnally and yellow nocturnally (versus a reddish brown dorsum with dark brown markings), pink (versus white to light brown) venter, indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold iris with limited, non-uniform black reticulations), dorsum with distinctive, white conical asperities (versus dorsum smooth) in males and eggs deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm (versus eggs in foam nest). From R. rhodopus Liu and Hu the new species differs by having a pink (versus yellowish) venter, indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold iris with limited, non-uniform black reticulations), dorsum with distinctive, white conical asperities (versus dorsum smooth) in males and eggs deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm (versus eggs in foam nest). From R. spelaeus Orlov, Gnophanxay, Phimminith, & Phomphoumy , the new species differs by having a pink venter (versus light gray venter covered by dark specks), indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold or yellowish cream iris with limited, non-uniform black reticulations), and dorsum with distinctive, white conical asperities (versus dorsum smooth) in males. From R. robertingeri Orlov, Poyarkov, Vassilieva, Ananjeva, Nguyen, Nguyen, & Geissler , Gracixalus lumarius sp. nov. differs by having a pink (versus white, grey or cream) venter, indistinct (versus distinct) tympanum and supratympanic fold, vomerine teeth absent (versus present), dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus bright gold or yellowish cream iris with limited, non-uniform black reticulations), dorsum with distinctive, white conical asperities (versus dorsum smooth) in males and eggs deposited as a tightly spaced array of nonpendent eggs on the wall of a phyptotelm (versus eggs in foam nest). Lastly, from Theloderma ryabovi Orlov, Dutta, Ghate, & Kent , the new species differs by having a brown dorsum diurnally and yellow nocturnally (versus light beige with lilac patterns and black spots), pink (versus dark grey) venter, dark gold iris with a dense, relatively uniformly distributed network of black reticulations (versus dark brown iris), and dorsum with distinctive, white conical asperities (versus dorsum with only small dorsal asperities) in males.
Molecular data. Our preliminary molecular analysis supports our placement of G. lumarius sp. nov. in the genus Gracixalus (1.00 Bayesian posterior probability; Fig. 8 View FIGURE 8 ). G. lumarius sp. nov. falls within “Clade I” of Gracixalus ( Rowley et al. 2011) , which also includes the type specimen of the genus, G. gracilipes (1.00 Bayesian posterior probability). Tree toplogy was near-identical when we excluded pairwise comparisons for gapped sites.
Molecular differentiation among G. lumarius sp. nov. and all other Gracixalus species for which comparable molecular data is available, is high, with 13.3–15.91% uncorrected sequence divergence at the 16S rRNA gene fragment examined. This degree of pairwise divergence in the 16S rRNA gene in frogs is strongly indicative of differentiation at the species level ( Vences et al. 2005). Our preliminary molecular analysis recovers the same relationships within the genus Gracixalus as Rowley et al. 2011 (eg. ‘Clade I’ and ‘Clade II’), but more extensive molecular analysis will be required to resolve the evolutionary relationships within the genus and among the family Rhacophoridae .




| T |
Tavera, Department of Geology and Geophysics |
| V |
Royal British Columbia Museum - Herbarium |
| UNS |
University of Science, Ho Chi Minh City, Vietnam |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |