
Lasianobia nainysi Saldaitis, Volynkin & Zahiri, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4927.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:F34255DE-60F9-4A66-AEEE-DC43635E786A |
|
DOI |
https://doi.org/10.5281/zenodo.4549235 |
|
persistent identifier |
https://treatment.plazi.org/id/624F5B08-6A2F-FFB8-A295-F184FE513276 |
|
treatment provided by |
Plazi |
|
scientific name |
Lasianobia nainysi Saldaitis, Volynkin & Zahiri |
| status |
sp. nov. |
Lasianobia nainysi Saldaitis, Volynkin & Zahiri , sp. nov.
( Figs 1–4 View FIGURES 1–8 , 9, 10 View FIGURES 9–12 , 15 View FIGURES 13–16 )
http://zoobank.org:act: 1951DE57-90DE-485A-8A9C-317509E5FBFC
Type material. Holotype ( Figs 1 View FIGURES 1–8 , 9 View FIGURES 9–12 ): male, “ China, N.W. Sichuan, Cho La Shan, Cho La pass, H- 4700m, N31°56’35’’ E98°55’56’’, 05.vii.2019, Butvila & Saldaitis leg.”, gen. slide No.: AV5614 (Coll. ASV / WIGJ). GoogleMaps
Paratypes ( Figs 2, 3 View FIGURES 1–8 , 10 View FIGURES 9–12 ): 13 males, the same data as for the holotype, gen. slide Nos.: AV5583, AV5615 (Colls AFM & ASV) GoogleMaps ; 40 males, 1 female, China, N.W. Sichuan, Cho La Shan, Cho La pass, 4450m, N31°55’07 E98°57’11’’, 01.vii.2019, Butvila & Saldaitis, gen. slide No.: AV5856 (female) (Colls AFM, ASV, CeNak, PGM/ HNHM & WIGJ) GoogleMaps ; 5 males, China, Sichuan, 50 km NW of Manigango city, 4300m, 13–16.vii.2005, gen. slide Nos.: GYP 2273, 5343 (Coll. PGM) .
Etymology. The species is named after Mr Saulius Nainys (Kriukai, Lithuania) for his enthusiastic Lithuanian entomology support.
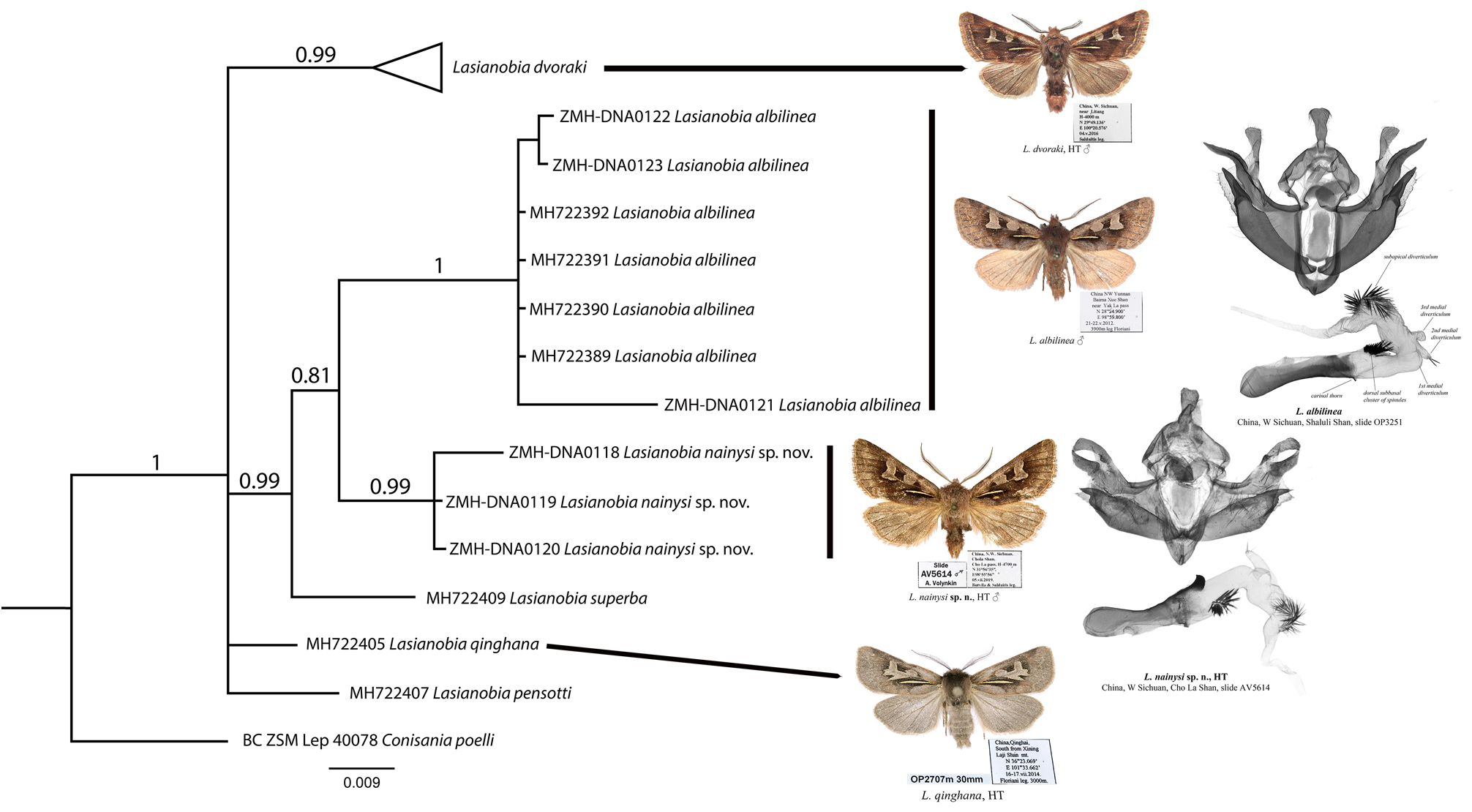
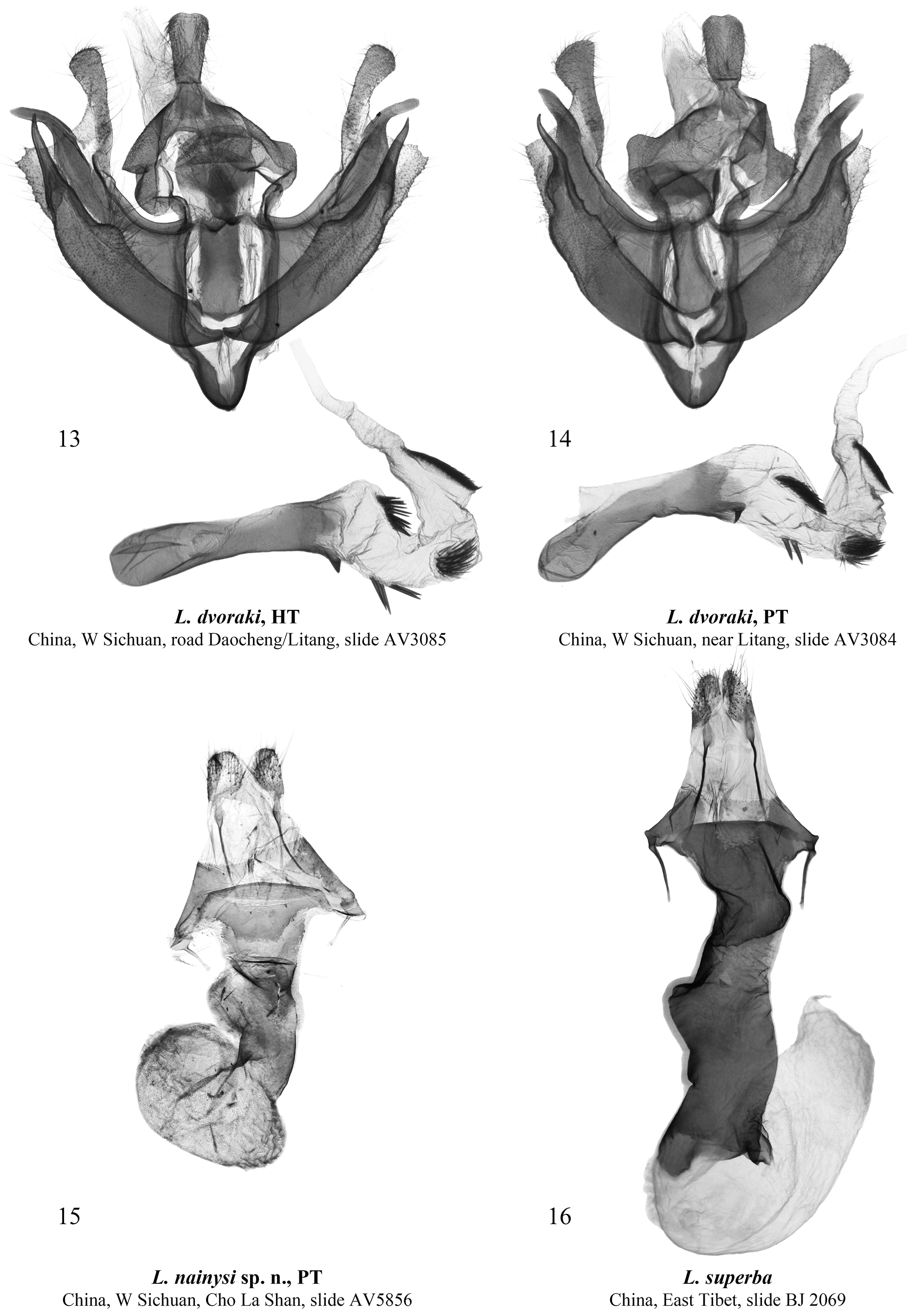
Diagnosis. Lasianobia nainysi sp. nov. ( Figs 1–4 View FIGURES 1–8 ) is most similar externally to L. albilinea ( Figs 5, 6 View FIGURES 1–8 ), but distinguishable by its more elongate forewing apex, slightly darker coloration of both wings and narrower reniform stigma. Additionally, the subbasal longitudinal dash of the new species is somewhat more elongate than in L. albilinea . The new species is also reminiscent of L. dvoraki ( Figs 7, 8 View FIGURES 1–8 ), but L. nainysi is somewhat larger, its coloration is darker, the reniform stigma has rounded posterior corners (that has pointed corners in L. albilinea ) and the subterminal line is more diffuse. The male genitalia of L. nainysi ( Figs 9, 10 View FIGURES 9–12 ) are similar to those of L. albilinea ( Figs 11, 12 View FIGURES 9–12 ), but differ in the somewhat less elongate costal process, the markedly larger carinal thorn of the aedeagus, and the slightly shorter cluster of spines on the subapical diverticulum. The male genital capsule of the new species has the wider costal process with a pointed tip, whereas in L. dvoraki it is narrower and bearing a rounded tip ( Figs 13, 14 View FIGURES 13–16 ). In the aedeagus of the new species, the carinal process is more robust than that of L. dvoraki . The vesica of L. nainysi lacks a ventral subbasal cluster of spinules (it is present in L. dvoraki ) but has a 3 rd medial diverticulum, which is absent in L. dvoraki . The 2 nd medial diverticulum of the new species bears only few spinules versus a wide cluster of numerous spinules in L. dvoraki . Additionally, the subapical diverticulum of L. nainysi bears noticeably wider and more elongate cluster of spinules than that of L. dvoraki . Females of L. albilinea and L. dvoraki are unknown therefore it is not possible to compare the female genitalia of the new species ( Fig. 15 View FIGURES 13–16 ) with those of L. superba ( Fig. 16 View FIGURES 13–16 ) only. The ductus bursae of L. nainysi is conspicuously shorter, narrower and more weakly sclerotized than that of L. superba . The corpus bursae is narrower, and the appendix bursae is markedly less elongate compared to those of L. superba . Additionally, the apophyses anteriores and posteriores of L. nainysi are less elongate and narrower than those of L. superba .
Molecular data. DNA barcode data from specimens of L. nainysi are significantly divergent from those of L. albilinea (2.24%), L. superba (2.08%), L. qinghana (2.08%) and L. dvoraki (2.25%). The tree topology generated by the BI analysis ( Fig. 17 View FIGURE 17 ) was very similar to the NJ-tree confirming the close association (PP = 0.81) between the new species (ZMH-DNA0118, ZMH-DNA0119, and ZMH-DNA0120) and a cluster of seven specimens of L. albilinea from China ( Fig. 17 View FIGURE 17 ). Comparison of DNA sequence data of the new species with its potential sister species ( L. albilinea ) revealed 14 polymorphic sites (single nucleotide polymorphisms, SNP) ( Table 2 View TABLE 2 ). Only two out of 14 base substitutions occur in the first position of the codon and the rest (12 sites) occur in the third codon positions ( Table 2 View TABLE 2 ). Only one mutation at position 376 was a non-synonymous substitution that changed the amino acid from AGC (Serine) to AGA (Arginine). Relationships inferred from DNA barcode data should be treated with caution given their length (658 bp), and are likely to change with addition of more molecular data and expanding taxon sampling.
Description. Adult ( Figs 1–4 View FIGURES 1–8 ). Forewing length 14–17.5 mm in males (17 mm in holotype) and 17 mm in females. Male antenna shortly bipectinate, female antenna filiform. Coloration of body and forewings reddish brown. Basal two thirds of forewing with intense pale pinkish brown suffusion along the costa. Antemedial and postmedial lines indistinct, thin, brown, smoothly curved. Subbasal longitudinal dash thin, pale yellowish, outlined with fine black scales, somewhat curved medially. Subterminal line indistinct, thin, pale, angled at apex. Veins in the terminal area weakly suffused with blackish scales. Orbicular stigma pale ochreous brownish, elliptical, its anterior and posterior margins sometimes diffuse. Reniform stigma pale ochreous-brown, with short extensions along the cubital veins. Forewing cilia brown. Hindwing dark brownish-grey, slightly lighter basally, with dark suffusion along the veins. Discal spot thin, semilunar, diffuse. Hindwing cilia dark greyish-brown.
Male genitalia ( Figs 9, 10 View FIGURES 9–12 ). Uncus dorso-ventrally flattened, broad, trapezoidal, with rounded distal corners, weakly setose. Tegumen relatively short but broad. Penicular lobes broad, ear-shaped. Juxta elongate, conical in shape. Vinculum V-shaped, rounded at base. Transtilla broad, lobe-like. Valves symmetrical, evenly curved medially. Costa of valva heavily sclerotized, its dorsal margin evenly curved and heavily sclerotized. Costal process robust, slightly curved medially, narrowed distally and pointed apically. Cucullus narrow, triangular with rounded corners, weakly setose; neck of cucullus elongate and narrow. Sacculus broad, with short, round, medial protrusion on its dorsal surface. Distal saccular process robust, elongate, peak-shaped, with wide basal plate. Clasper short, triangular, without processes. Aedeagus relatively short but broad, cylindrical in shape. Aedeagus carina with large and heavily sclerotized triangular thorn. Vesica tube-like, curved dorsally in its medial section; its subbasal section with short band-like cluster of numerous robust dorsal spinules. 1 st medial diverticulum of vesica broad but very short, not prominent. 2 nd medial diverticulum small, conical, bearing a few spinules. 3 rd medial diverticulum short and narrow, without spinules. Subapical diverticulum broad and short, armed with broad and elongate cluster of numerous spinules.
Female genitalia ( Fig. 15 View FIGURES 13–16 ). Ovipositor short, broadly conical. Papilla analis trapezoidal with rounded corners, weakly setose. Apophyses elongate and thin, apophyses posteriores ca. 2 times longer than apophyses anteriores. Ostium bursae wide. Antrum broad, funnel-shaped, heavily sclerotized. Anterior section of ductus bursae elongate, dorso-ventrally flattened, with broad and rounded lateral protrusion on its left side. Corpus bursae short and narrow, densely scobinated, with gelatinous right lateral margin. Appendix bursae broader than corpus bursae, laterally positioned, densely scobinate.
Distribution and bionomics. Series of males and single female were collected at light in first part of July in a remote part of west China Sichuan province at Cho La pass near the Maniganggo. The new species was collected at elevations ranging from 4300 to 4700 meters in alpine mountain zone dominated by various herbaceous vegetation and scarce shrubs ( Figs 18–19 View FIGURES 18–19 ).



| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
| PGM |
Pacific Grove Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Noctuinae |
|
Genus |