
Diphylla ecaudata SPIX, 1823
|
publication ID |
https://doi.org/ 10.1093/mspecies/sead003 |
|
persistent identifier |
https://treatment.plazi.org/id/61778794-FF8D-2E37-FC15-C411FF64FA97 |
|
treatment provided by |
Plazi |
|
scientific name |
Diphylla ecaudata SPIX, 1823 |
| status |
|
Diphylla ecaudata SPIX, 1823 View in CoL
Hairy-legged Vampire Bat
Diphylla ecaudata Spix, 1823:68 View in CoL . Type locality “ Brazil;” restricted to Rio San Francisco, Bahia by Cabrera (1958:94).
Gl [ossophaga]. diphylla J. B. Fischer, 1829:133. Type locality “Brasilia;” based on Diphylla ecaudata Spix, 1823 View in CoL .
Diphylla centralis O. Thomas, 1903:378 . Type locality “Boquete,” Chiriqui, Panama.
Diphylla ecaudata centralis View in CoL : Felten, 1956a:364. Name combination.
CONTEXT AND CONTENT. Content same as for genus. Whether or not Diphylla ecaudata View in CoL is monotypic is unresolved in the literature. Ojasti and Linares (1971), Jones and Carter (1976), and Greenhall et al. (1984) recognized two subspecies, Diphylla ecaudata centralis ( Thomas, 1903) View in CoL distributed in Mexico and Central America and Diphylla ecaudata ecaudata ( Spix, 1823) View in CoL distributed in South America. Burt and Stirton (1961), Hall (1981), and Koopman (1988) considered D. ecaudata View in CoL as monotypic and referred to these subspecies as morphologically indistinguishable. More recently Simmons (2005) considered the species monotypic, whereas Aguirre (2019) and Burgin et al. (2020) recognized the two previously mentioned subspecies. It has been suggested that D. ecaudata View in CoL needs to be revised ( Kwon and Gardner 2008).
NOMENCLATURAL NOTES. The generic name derives from the Latin word di - two + - phylla from the Greek word “phyllon” which means leaf and refers to the bifoliate nose leaf (thus the specific epithet, diphylla). The specific name ecaudata is related to the absence of a tail and interfemoral membrane ( Spix 1823). Common name in Spanish is vampiro de patas peludas ( Mexico — Wilson et al. 1985), and in Portuguese it is vampiro de pernas peludas ( Brazil — Aguiar 2007).
DIAGNOSIS
Diphylla ecaudata ( Fig. 1 View Fig ) is a medium-sized bat similar to the other species of Desmodontinae , the common vampire bat ( Desmodus rotundus ) and the white-winged vampire bat ( Diaemus youngii ). Diphylla ecaudata can be differentiated by its short and wide ears (wider than long) compared to Diaemus and Desmodus with their longer and more pointed ears (clearly longer than wide). Diphylla ecaudata differs from Diaemus and Desmodus in its well-developed secondary noseleaf, located behind the primary noseleaf ( Fig. 2 View Fig ; Bhatnagar 1988; Koopman 1988; Kwon and Gardner 2008; Medellín et al. 2008; dos Santos and Lopes 2015; Scheffer et al. 2015; Álvarez-Castañeda et al. 2017).
Wing tips in D. ecaudata and Desmodus are dark and lack white spots, whereas Diaemus presents conspicuous white wing tips. Legs are thickly haired in D. ecaudata , whereas Desmodus and Diaemus have nonhirsute legs. The edge of the interfemoral membrane in D. ecaudata is covered with fur, very narrow, and inserted in the middle of the calcar, leaving the end of the calcar free (digitiform). In Diaemus and Desmodus the interfemoral membrane is naked or has little fur, and is wider; the calcar is vestigial in Desmodus and absent in Diaemus ( Greenhall and Schutt 1996) . The thumbs (pollices) in D. ecaudata and Diaemus measure one-eighth the length of the third digit, and there is a single metacarpal cushion, in contrast with the thumbs of Desmodus , which are longer, measuring one-fifth the length of the third digit, and have two metacarpal cushions ( Schutt and Simmons 2006). Diphylla ecaudata features a metacarpal without pads, unlike Diaemus with a single pad under the metacarpal, and Desmodus with two pads ( Koopman 1988; Scheffer et al. 2015).
The dental formula of D. ecaudata is i 2/2, c 1/1, p 1/2, m 2/2, total 26, whereas the total number of teeth is 22 (i 1/2, c 1/1, p 1/2, m 2/1) in Diaemus and 20 (i 1/2, c 1/1, p 1/2, m 1/1) in Desmodus ( Medellín et al. 2008; Álvarez et al. 2017). The lower incisors are broad, flat, complex, and larger than those in Desmodus and Diaemus ( Greenhall et al. 1984; Koopman 1988; Davis et al. 2010; Scheffer et al. 2015). In D. ecaudata , the lower incisors are arranged in a continuous crest of small beadshaped lobes, whereas a diastema separates the central incisors in Desmodus and Diaemus ( Davis et al. 2010) . The number of lobes (cusps) in the lower incisors of D. ecaudata is variable; four lobes have been described in the medial incisor, and 5–8 in the lateral incisor, with 6 being the most common. In contrast, Desmodus and Diaemus have bilobed lower incisors ( Bhatnagar et al. 1992, Davis et al. 2010; see “Form and Function”).
Diphylla ecaudata has a longitudinal palate groove ( Uieda 1986), as opposed to Desmodus and Diaemus , which lack this structure. Instead, they have longitudinal grooves on the ventral surface of their tongue, a prominent groove on their lower lips, and another in the diastema between the inferior central incisives. Diphylla ecaudata presents a fully fused mandibular symphysis, versus an unfused symphysis in Desmodus and Diaemus ( Davis et al. 2010) .
GENERAL CHARACTERS
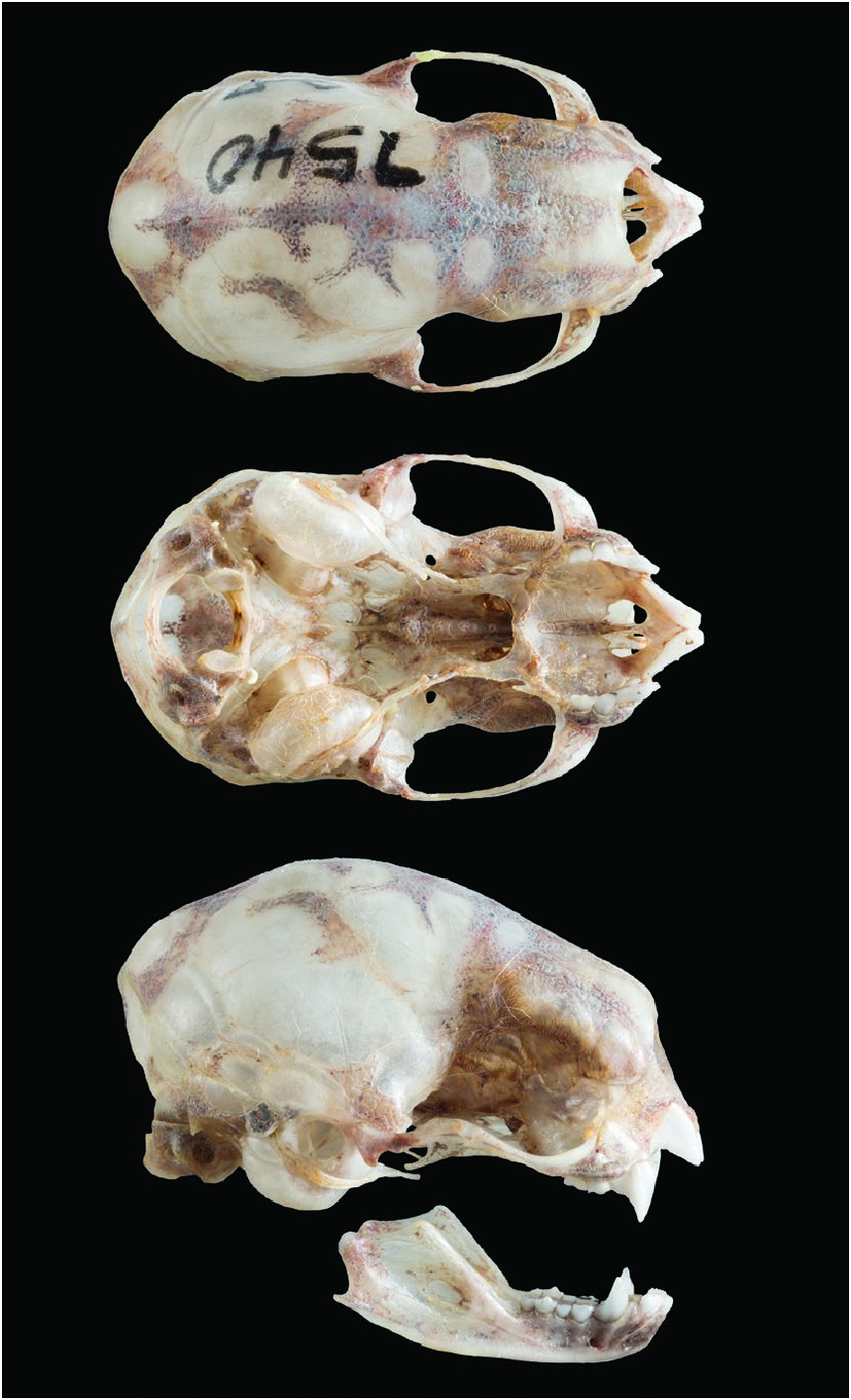
The cranium is characterized by being tall and wide, lacking a postorbital process ( Fig. 3 View Fig ). There is an ample interorbit area and an interpterygoid space with almost parallel sides. The coronoid apophysis is low but is elevated over the condyle (Model 1; Greenhall et al. 1984; Bhatnagar 1988; Koopman 1988; Scheffer et al. 2015). Greatest cranium length is 23.5 mm ( Husson 1962). Males and females are similar in size. Mean head–body length (mm) in adults of both sexes was: 78.60 (n = 43— Elizalde-Arellano 2003). Mean forearm lengths (mm; ± SD; parenthetical n) were: sexes combined, 52.37 ± 2.19 (21); females only 52.90 ± 2.20 (16); and males only 52.66 ± 2.22 (5— Patricio-Costa 2015). Body mass (g) varies according to feeding status. Prior to feeding, adults had a mean body mass of 28.5 (range 20–35; n = 26), whereas mean body mass of postfeeding adults was 32.7 (n = 14, range 25–41— Elizalde-Arellano 2003). Diphylla ecaudata has small, ample, rounded ears. The interfemoral membrane, limbs, and ears are densely furred. It has a short thumb with no cushion under the metacarpal. Diphylla ecaudata has large bright eyes, a naked tragus, and lacks a tail.
DISTRIBUTION
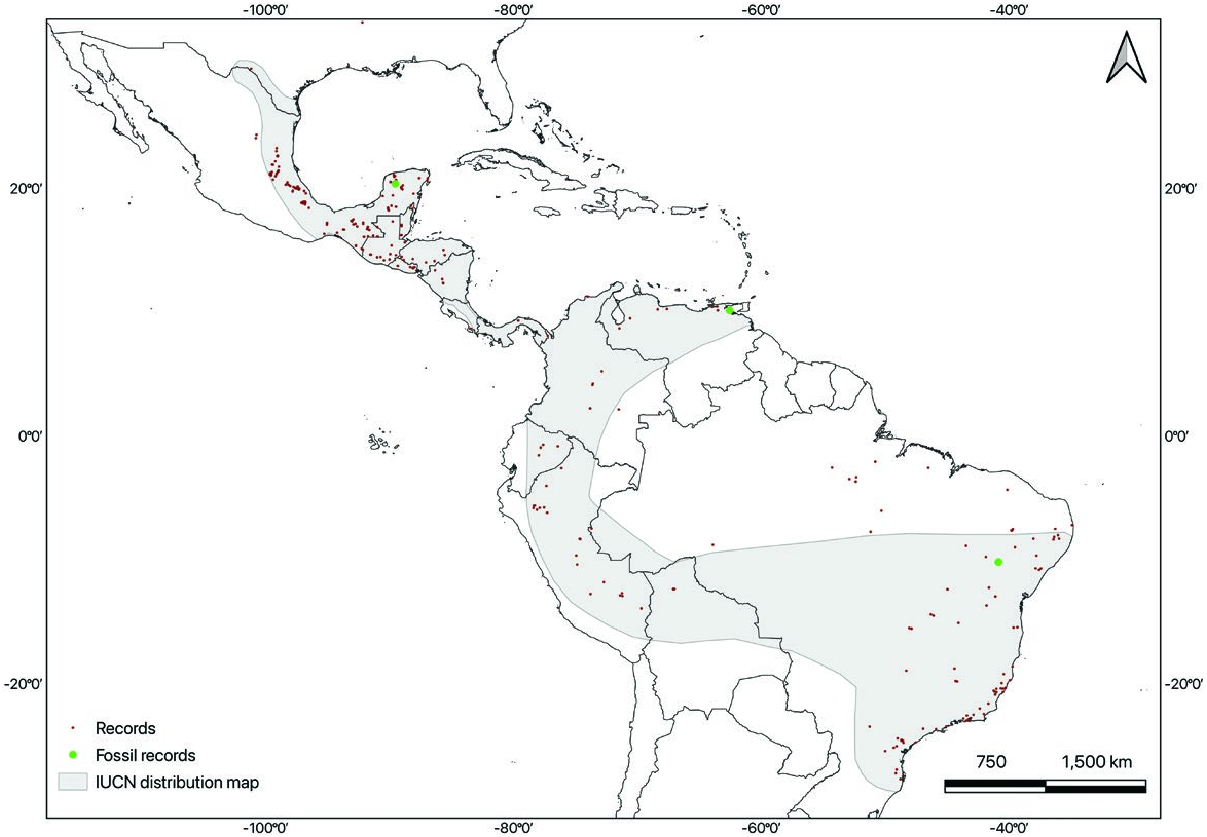
Diphylla ecaudata is geographically restricted to the Americas ( Fig. 4 View Fig ). It can be found from eastern Mexico through Central America and well into South America ( GBIF 2021). There is a single report of a female in south Texas, United States ( Reddell 1968; Simmons 2005), but it is likely an instance of an accidental migrant. In Mexico, it can be found from southern Tamaulipas ( Elizalde-Arellano et al. 2007; Ceballos and Arroyo 2012) and San Luis Potosí ( Wilson et al. 1985), southward through part of Guanajuato ( Magaña-Cota et al. 2010), Puebla ( Vargas-Miranda et al. 2008), Veracruz ( Martínez-Gallardo and Sánchez-Cordero 1997; Coates et al. 2017), Oaxaca ( Briones-Salas et al. 2015), Chiapas ( Horvath et al. 2001; Cruz-Lara et al. 2004; Escobedo et al. 2005; Lorenzo et al. 2017), Yucatán ( Arita 1997), and Quintana Roo ( Pozo de la and Escobedo 1999). It is distributed continuously throughout Central America in Guatemala, Belize, El Salvador, Honduras, Nicaragua, Costa Rica, and Panama ( Dalquest and Hall 1947; Jones 1966; McCarthy 1987; Sampaio et al. 2016), and into South America, with records in Venezuela ( Ojasti and Linares 1971; Handley 1976), Colombia ( Aellen 1970; Castro 2016; Echavarría et al. 2017), Ecuador ( Albuja, 1983), Bolivia ( Siles et al. 2003), Perú ( Tuttle 1970; Graham and Barkley 1984; Hutterer et al. 1995; Solari et al. 2001; Quintana and Pacheco 2007), and Brazil, where there are reports of its presence in 20 states, including the Amazon State and the Federal District ( Esbérard et al. 2005; Faria et al. 2006; Feijó and Langguth 2011; Rocha et al. 2014; Santos and Lopes 2015; see Supplementary Data SD1). It occurs from sea level up to about 1,900 m ( Sampaio et al. 2016).
FOSSIL RECORD
The first late Pleistocene record (a humerus) of Diphylla ecaudata in Mexico was reported from Loltún Grutas (20°24ʹN, 89°32ʹW) in the state of Yucatan ( Álvarez 1982). Additional fossil and subfossil remains of late Pleistocene–Holocene age individuals were subsequently identified from several deposits within the caves ( Arroyo-Cabrales and Álvarez 1990; Arroyo-Cabrales et al. 2005). In Venezuela, there is a Quaternary fossil record from the El Guácharo Cave in the state of Monagas (10°l0ʹ27″N, 62°33ʹ07″W — Linares 1968). Late Pleistocene fossils of D. ecaudata have been found in the Toca da Boa Vista and Toca da Barriguda caves in the state of Bahia, Brazil ( Czaplewski and Cartelle 1998; Salles et al. 2014).
Model 1. — Exterior skull anatomy of Diphylla ecaudata ( ECOSUR [El Colegio de la Frontera Sur, San Cristóbal de Las Casas Unit , Chiapas, México Mastozoological collection] 7540) from Paraje Chojolhó, Chiapas, México. https://sketchfab.com/3d-models/ diphylla-ecaudata-exterior-skull-anatomy-62aba50945da43e394ee082 319a80194 .
FORM AND FUNCTION
Diphylla ecaudata possesses a more specialized dentition compared to the other hematophagous bats. Its upper incisors are shaped like a sickle ( Davis et al. 2010), lower incisors are large, possess between four and eight lobes, and lack a diastema ( Bhatnagar et al. 1992); upper canines are also large, though smaller than the upper incisors and sharpened as a result of wear by tooth-on-tooth friction against lower canines.
The upper incisors are less procumbent (61.02° ± 9.02 SD) than those of Desmodus (73.51° ± 3.92 SD) and Diaemus (76.21° ± 7.39 SD; n = 5 males and 5 females for Desmodus , n = 3 males and 3 females for Diaemus and Diphylla — Davis et al. 2010). The smaller angle of incisor procumbency found in D. ecaudata is associated with deeper and larger mandibular pits (cuplike pits in the mandible behind the lower incisors, where the tips of the upper incisors rest). The mean mandibular pit volume (mm 3) is 1.11 ± 0.19 SD in D. ecaudata , 0.79 ± 0.15 SD in Desmodus , and 0.58 ± 0.14 SD in Diaemus ; n = 5 males and 5 females for each of the three species ( Davis et al. 2010). The mean mandibular pit depth (mm) is 1.10 ± 0.14 SD in D. ecaudata , 0.82 ± 0.09 SD in Desmodus , and 0.68 ± 0.11 SD in Diaemus . Larger pits in D. ecaudata are coupled with a reduced bone in the anterior region of the mandible; the mean mandible length (mm) in D. ecaudata is 13.2 ± 0.2 SD, whereas in Desmodus it is 14.8 ± 0.5 SD and in Diaemus , 14.9 ± 0.3 SD. Not only do the pits occupy more space, but D. ecaudata also has a short symphysis (mean symphysis height in mm of D. ecaudata : 1.84 ± 0.11 SD, Desmodus : 1.70 ± 0.07 SD, Diaemus : 1.95 ± 0.07 SD) and a small symphyseal surface area (mean symphyseal surface area in mm 2 of D. ecaudata : 2.8 ± 0.04 SD, Desmodus : 3.9 ± 0.04 SD, Diaemus : 4.0 ± 0.04 SD; sample sizes as given above— Davis et al. 2010). It has been suggested that these differences in mandibular morphology and the previously described differences in the lower incisors (see “Diagnosis”) of the Desmodontinae are associated with differences in preferred prey ( Birney and Timm 1975; Davis et al. 2010). Diphylla ecaudata preferentially feeds on avian prey, whereas Diaemus youngii prefers avian blood but will feed on mammalian blood, and Desmodus rotundus feeds primarily on mammalian blood. The unique features present in D. ecaudata may be the result of adaptation facilitating its grip on bird skin which differs in thickness and texture in comparison to mammalian skin ( Birney and Timm 1975). Postcanine dentition is reduced both in number and size and contains minute nonocclusive premolars and molars shaped like razors ( Davis et al. 2010). Qualitatively, the mandible of D. ecaudata is gracile; the coronoid and anterior mandible are thin enough to appear particularly radiolucent ( Greenhall et al. 1984; Davis et al. 2010).
The large foramen magnum accounts for 40% of the greatest width of skull. Vertebral formula is 7C, 12T, and 5L, total 24; sacral vertebrae are fused and there are no caudal vertebrae. Sternum consists of the manubrium, six sternebrae, and the xiphoid process; the sternebrae are crested and fused ( Bhatnagar 1988).
The humerus presents a relatively prominent pectoral ridge and deep trochlea. The ulna is fused distally with the radius at about the middle of the radial shaft. At the pelvis, the pubic spine is long, diverging at the tip cranially; ischial rami are slender ( Bhatnagar 1988). The femur is thin and the fibula is incomplete ( Schutt and Simmons 2006). The calcar is large and digitiform. It acts as a sixth finger, which serves as an auxiliary grip for perching on tree branches ( Schutt and Altenbach 1997). Foot length is 11– 6 mm and tibia length is 19.9–23 mm; n = 3 males and 2 females ( Bhatnagar 1988).
The fur is long and soft, light to dark brown on the dorsum and lighter in the ventral region ( Scheffer et al. 2015). Hair length, measured in one individual, is 12 mm with a diameter of 22.1 µm ( Benedict 1957). Individual hairs lack a prominent basal bulb; melanin granules are distributed sparsely across the filament. No difference exists between the over and under fur ( Bhatnagar 1988).
Brain weight of D. ecaudata is 0.805 g (n = 9— Stephan et al. 1981). The volume of the pineal gland is 0.2302 mm 3 (n = 3— Bhatnagar et al. 1986). Dimensions of the main olfactory bulb in a single male specimen are length 2.90 mm and width 2.20 mm; the length of the accessory olfactory bulb is 200 µm, and width is 310 µm ( Frahm and Bhatnagar 1980), and its volume is 0.0457 mm 3 ( Frahm 1981). The vomeronasal organ is 1.54 mm long ( Bhatnagar 1988).
Diphylla ecaudata has a tongue with the typical histologic structures of mammals ( Schultz 1965), and despite its highly selective diet, which might make taste perception irrelevant, well-developed taste buds are present ( Obregón 2010). Genes for bitter flavor receptors, common in other bats, were reduced in the evolution of Desmodontinae ( Hong and Zhao 2014). In D. ecaudata , the cheek lacks sweat glands in the skin. The lips are structured longitudinally like a vasculo-nervous plexus, surrounding a structure that appears to be sensory. The esophagus contains three regions: cervical, thoracic, and abdominal. The first two portions have skeletal muscle fibers in the muscular tunic ( Obregón 2010), allowing the altruistic regurgitation habit that D. ecaudata displays in colonies ( Elizalde-Arellano et al. 2007). This muscular tunic transitions to smooth muscle in the abdomen, with a stomach-like columnar epithelium; its mucosae form short, thick projections into the lumen, not visible in the former portions. The transition from stratified squamous epithelium to simple columnar epithelium is gradual and occurs at the esophageal hiatus of the diaphragm. The stomach is shaped like a bagpipe with small, rounded projections on its outer surface which appear as a series of incomplete subcompartments on the inside. The gastric mucosa has elongated projections, suggesting a high capacity for expansion and storage. Cardiac and pyloric mucosae are similar, with scant, long projections, whereas fundic mucosae are uniform and median in length. Due to the pronounced bagpipe shape of the stomach, the cardia and pylorus are very close, particularly when the stomach is empty ( Obregón 2010).
A microscopic comparison of the small and large intestines of D. ecaudata and a frugivore species, Artibeus planirostris (flat-faced fruit-eating bat), revealed morphological differences that were most likely associated with the differences in diets between the two species ( Silva et al. 2020). Due to the nature of its diet, mostly proteinaceous and semifluid, the intestinal walls of D. ecaudata are less morphologically complex and specialized than those of the frugivore ( Silva et al. 2020). Intestines have a short relative length with an average of 18.04 ± 2.91 cm (± SD) from the pylorus to the end of the rectum (n = 4 males and 1 female — Silva et al. 2020).
All three portions of the small intestine of D. ecaudata are similar, it is macroscopically impossible to differentiate between the jejunum and ileum, and few histological differences help to distinguish them ( Obregón 2010; Silva et al. 2020). The cecum is absent. Evidence of an active immune system is suggested by a large amount of diffuse lymphoid tissue and solitary lymph nodes throughout the gastrointestinal tract. All three types of salivary glands, parotid, submandibular, and sublingual, are present. The liver is histologically similar to that of other mammals, with abundant hepatocytes, suggesting an excellent and rapid storage capacity ( Obregón 2010). The pancreas is in the abdominal cavity; the right lobe is close to and parallel to the descending duodenum, and the left lobe transversely extends caudally to the stomach. The mean mass of the pancreas is 0.1 g (n = 6; mean body mass = 24.4 g — Machado-Santos et al. 2013). It is covered by a thin capsule of loose connective tissue that extends into the organ forming a series of septa, which subdivide the gland into visibly distinct lobes. It consists of an exocrine portion with acini and a duct system starting with small central acinar cells that lead to the intralobe ducts covered by a simple squamous or cuboid epithelium, and an endocrine portion comprised of Islets of Langerhans. These islets are irregular in shape and easily identified by their eosinophilic staining properties. The average number (± SD) of Islets of Langerhans is 23,900 ± 9,770.54, with an average diameter of 56.94 µm, and a mass of about 0.48 mg, their volumetric density is 4.1% (n = 6 males — Machado-Santos et al. 2013). Insulin-immunoreactive (IR) cells occur in the middle of the islets, and glucagon-IR and somatostatin-IR cells are located peripherally; somatostatinsecreting cells are also found in isolation. As for the volume density of endocrine cells, it is larger for β cells (54.8%) and then decreases for α cells (16.2%) and δ cells (14.3%— Machado-Santos et al. 2013).
Ovaries are oval-shaped, and average 1.5 mm (0.7–2.0 mm) long and 1.3 mm (0.3–2.2 mm) wide (n = 18— Elizalde-Arellano 2003). Oviducts (called “uterine tubes” in anatomic literature) are short and contoured around the ovaries. The uterus is bicornuate and symmetrical, with thin uterine horns in nonpregnant individuals but these enlarge noticeably during early gestation. A uterine horn with implanted fetuses can distend up to 27 times its original size as the fetus develops, whereas the nonimplanted uterine horn becomes imperceptible. Females are monotocous ( Elizalde-Arellano 2003).
The penis has an average length of 12.2 mm (9.5–14.1; n = 12), lacks a baculum, and is covered with a thick foreskin with numerous folds ( Elizalde-Arellano 2003). Accessory sex glands such as bulbourethral (formerly called, “Cowper’s glands”) and seminal vesicles have not been reported in this species ( Brown et al. 1971; Elizalde-Arellano 2003).
Males have ovoid testicles with an average length of 4 mm (2.8–5.0 mm) and width of 3.4 mm (2.6–3.8 mm; n = 12— Elizalde-Arellano 2003). Position and size of the testicle are related to age: newborns and juveniles have abdominally positioned testicles, subadults have scrotal or inguinal testicles, whereas adults, after 231 days of age, have permanent scrotal testicles throughout the year, indicating they are sexually active continuously ( Delpietro and Russo 2002; Elizalde-Arellano 2003). The testicles and epididymides are joined longitudinally and have a highly related degree of development. The cauda of each epididymis is continuous with the deferent duct and joins the dorsal part of the prostate. The ventral part of the prostate joins with the bladder and the distal part with the urethra ( Elizalde-Arellano 2003).
As for testicular characterization and spermatogenesis, the species has a gonadosomatic index (%) of 0.49 ± 0.17 (± SD), a tubulesomatic index (%) of 0.47 ± 0.16 (± SD), and the length of the seminiferous tubules per gram of testis (m/g) is 32.20 ± 2.00 (± SD; n = 6— Silva et al. 2019). The frequency of phases within the seminiferous epithelium cycle is 56.20% premeiotic phase, 9.30% meiotic phase, and 34.50% in the postmeiotic phase. Its testicular pattern includes common characteristics present in other bat species and is similar to that of other mammals. Diphylla ecaudata has a notably high efficiency of Sertoli cells, which are adept at sustaining germinal cells, and it has one of the highest sperm production rates of all bat species ( Silva 2018; Silva et al. 2019). Mean measurements (µm) ± SD characterizing spermatozoa are head length 4.57 ± 0.160, head width 3.46 ± 0.126, acrosome length 2.89 ± 0.154, nuclear length 4.22 ± 0.154, and medium piece 9.60 ± 0.294 (n = 3— Forman and Genoways 1979).
Diphylla ecaudata is capable of postprandial thermoregulation. In this phase, oxygen uptake increases by 50%, and average body temperature (32.4°C) increases by around 2°C ( McNab 1969, 1973). It has a metabolic pattern characterized by small carbohydrate and lipid reserves during fasting, which makes it highly susceptible to food restriction in periods over 36 h, suggesting it cannot live over 24–36 h without food. Glucogenesis pathway deficiency has been suggested, so their hematophagous diet may be associated with a high susceptibility to food scarcity ( Dias-Gomes 2008).
ONTOGENY AND REPRODUCTION
Ontogeny.— The oocyte pellucid zone is lost after fertilization during its passage through the oviduct. Embryo implantation is ipsilateral and reaches amniogenesis within 3 months ( Elizalde-Arellano 2003). Gestation is palpable at 141 days. Newborn Diphylla ecaudata are precocious, with open eyes and bodies covered in fur; weight at birth ranges between 7.5 ( Delpietro and Russo 2002) and 9 g ( Elizalde-Arellano 2003), and mean forearm length (mm ± SD) is 26.2 ± 0.2 in males (n = 5) and 26.4 ± 1.17 in females (n = 6— Delpietro and Russo 2002). Upon birth, deciduous teeth with a dental formula of i 2/2, c 1/1, p 2/2, total 20 are present ( Birney and Timm 1975; Bhatnagar 1988). During their first 17 days, young strongly adhered to their mother’s nipples, with their eyes constantly open and wary of objects moving around them. Lactation is long, and weaning occurs after 217 days on average ( Delpietro and Russo 2002), which contrasts with most other bats (e.g., 23 days in Carollia perspicillata [Seba’s short-tailed bat], 25 in Myotis lucifugus [little brown myotis], 35 in Eptesicus fuscus [big brown bat], 66 in Artibeus jamaicensis [Jamaican fruit-eating bat]— Kunz and Hood 2000); however, it is similar to other vampire bats (278 days in Desmodus rotundus — Delpietro and Russo 2002). After weaning, young are fed by the mother by regurgitation of blood until they are 223 days old ( Delpietro and Russo 2002).
Young have slow postnatal growth ( Elizalde-Arellano 2003); they first fly at around 57 days of age, reach their adult weight in an average of 178 days, and epiphyseal–diaphyseal fusion of the metacarpal and phalangeal bones occurs at 215 days. Young males present scrotal testes at 231 days and distance themselves from the main colony at 235 days. The first gestation in females happens at 367 days ( Delpietro and Russo 2002).
Reproduction.— In general, hematophagous bats are thought to have a continuous polyestrous reproductive pattern ( Racey and Entwistle 2000); nonetheless, in Diphylla ecaudata some studies assume the existence of a reproductive season in Mexico ( Dalquest 1955) and Argentina ( Delpietro and Russo 2002), where 92% of births are recorded in spring–summer. Vaginal cytology has helped identify all four phases of the estrous cycle (proestrus, estrus, metestrus, and diestrus), although the duration of each is still unknown ( Elizalde-Arellano et al. 2008).
Female individuals are deemed sexually mature at 9 months old ( Scheffer et al. 2015). Delpietro and Russo (2002) suggest that the gestation period could last 151–166 days, and Elizalde-Arellano (2003) estimates it could last 5–7 months. Females bear offspring once a year ( Bhatnagar 1978; Schmidt 1988; Delpietro and Russo 2002; Elizalde-Arellano 2003; Elizalde-Arellano et al. 2008), although biyearly births have been reported in El Salvador ( Felten 1956b). There is one report of a breech birth ( Bhatnagar 1978). Lactating females who lose their young have been observed to continue feeding the young of other females. In these cases, adoptive mothers and biological mothers usually wean the young and get pregnant at the same time ( Delpietro and Russo 2002).


| ECOSUR |
El Colegio de la Frontera Sur (Mexico) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diphylla ecaudata SPIX, 1823
| Ocampo-González, Paola, Espinoza-Medinilla, Eduardo E., Rioja-Paradela, Tamara M. & López-Wilchis, Ricardo 2023 |
Diphylla ecaudata centralis
| Felten H. 1956: 364 |
Diphylla centralis O. Thomas, 1903:378
| Thomas O. 1903: 378 |
Diphylla ecaudata
| Cabrera A. 1958: 94 |
| Spix J. B. 1823: 68 |