
Wernerius mumai ( Sissom, 1993 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.4648725 |
|
publication LSID |
lsid:zoobank.org:pub:CBB9D2AA-980B-4F22-9BA2-C0BAF01F7F1 |
|
DOI |
https://doi.org/10.5281/zenodo.4773106 |
|
persistent identifier |
https://treatment.plazi.org/id/61280E24-0432-FFC4-97D7-069356DDFC92 |
|
treatment provided by |
Carolina |
|
scientific name |
Wernerius mumai ( Sissom, 1993 ) |
| status |
|
Wernerius mumai ( Sissom, 1993) View in CoL
( Figures 1−21 View Figures 1–2 View Figures 3–4 View Figures 5–11 View Figures 12–15 View Figures 16–17 View Figures 18–20 View Figure 21 , Tables 1−2 View Table 1 View Table 2 )
http://zoobank.org/urn:lsid:zoobank.org:act:6DC0F0B2- 2926-4BB9-84E9-AC062D2D83DF
Vaejovis mumai Sissom, 1993: 64–65 , 68, figs. 1−7; Sissom, 2000: 551.
Wernerius mumai: Soleglad & Fet, 2008 View in CoL : 88; Webber et al., 2012: 2.
MATERIAL EXAMINED (topotypes). USA: Arizona, Mohave County, Gold Road , 35.04119ºN 114.37191 Wº, 986 m a. s. l. ( Figs. 18–19 View Figures 18–20 ), 20 March 2018, 2♂, RA2907 ( USNM), RA2650 About USNM ( RFA), leg. R. F. Ayrey, 11 March 2012, 1♀, RA573, 1♂, RA574 ( RFA), 16 March 2012, 2♂, RA575, RA576 ( RFA), 24 March 2020, 1♀, RA593 ( RFA), leg. R. F. Ayrey & M. DeBoer-Ayrey, 04 April 2016, 1♂, RA2332 ( RFA), 1 ♀, RA2331 ( RFA), leg. R. F. Ayrey, 12 April 2020, 1♂, RA2924 ( RFA), 13 April 2020, 1♀, RA2923 ( RFA), 14 April 2020, 1♂, RA2928 ( RFA), 19 April 2020, 1♀, RA2927 ( RFA), leg. R. F. Ayrey & M. DeBoer-Ayrey .
DIAGNOSIS. Small (27.74 mm) scorpions. Color is reddish brown, lighter on the legs. (see Fig. 1 View Figures 1–2 ). Pedipalp movable finger with 7 ID denticles and fixed finger with 6. Carapace of male is longer than the fifth metasomal segment. Pectinal tooth count for males 12.31 [n=16]; for females 11.40 [n=10]. Prominent subaculear tubercle.
DESCRIPTION. Based on a topotype male (specimen RA2907), unless otherwise noted; see Fig. 4 View Figures 3–4 for dorsal and ventral views. Coloration ( Figs. 1–2 View Figures 1–2 ). Color is reddish brown, lighter on the legs.
Carapace ( Fig. 5 View Figures 5–11 ). Anterior margin of carapace moderately emarginated, posterior margin slightly emarginated. Carapace finely granular, with larger granules dispersed throughout. Three lateral eyes on each side. Six macrosetae situated along the anterior edge of carapace. One pair of macrosetae positioned behind lateral eyes on each side. One pair of macrosetae positioned directly behind median eyes, located on either side of median furrow. Median furrow moderate and traverses entire length of carapace. Ratio of median eyes location from anterior edge/carapace length 0.36; carapace length/width at median eyes 1.39. Carapace of male is longer than metasomal segment V.
Mesosoma. Tergites finely granular with vestigial median carina on tergites I−VI. Tergite VII with weak median carina and strong dorsal lateral and lateral supramedian granular carinae. Sternites III−VI finely granular and without carinae.
Sternite VII with granular ventral lateral carinae on posterior half. Presternites smooth. Spiracles ovoid with median side rotated 35 degrees from posterior sternite margin. Sternites with variable number of microsetae.
Sternum ( Fig. 6 View Figures 5–11 ). Sternum is type 2.
Genital Operculum ( Fig. 6 View Figures 5–11 ). Sclerites separated on posterior one-fifth.
Pectines ( Fig. 6 View Figures 5–11 ). Pectinal tooth counts 12/12 [n=5], 13/12 [n=1] and 13/13[n=2], with a mean of 12.31 [n=16], SD = 0.464 for males; 11/11 [n=3] and 12/12[n=2], with a mean of 11.40 [n=10], SD = 0.516 for females. All male pectinal teeth have exterodistal angling with large sensorial area. All female pectines have at least 1 and sometimes 2 of the most proximal teeth smaller and lacking sensorial area, mean of 1.10 [n=10]. Middle lamellae 8/8. Fulcra are present. Each fulcra with 1−3 central setae.
Chelicerae. Dorsal edge of movable cheliceral finger with two subdistal (sd) denticles. Ventral edge is smooth, with well-developed serrula on distal half.
Pedipalps ( Figs. 10 View Figures 5–11 , 21 View Figure 21 ). Trichobothrial pattern type C ( Vachon, 1974). Trichobothria ib/it at base of fixed finger. Pedipalp ratios: chela length/width 3.31; femur length/width 2.97; patella length/width 2.83; fixed finger length/carapace length 0.58.
Chela . External carinae weak; D1 and D3 carinae weak. D4 and D5 carinae weak, granular, with individual granule size increasing distally. Internal and ventral carinae weak to obsolete. Fixed finger median (MD) denticles aligned and divided into 6 subrows by 5 outer (OD) denticles and 6 ID denticles. Movable finger with 6 subrows, 5 OD denticles and 7 ID denticles (Soleglad & Sissom, 2001).
Femur. Carinae moderate.
Patella. Carinae strong, internal surface with very large granules on the DPSc carina.
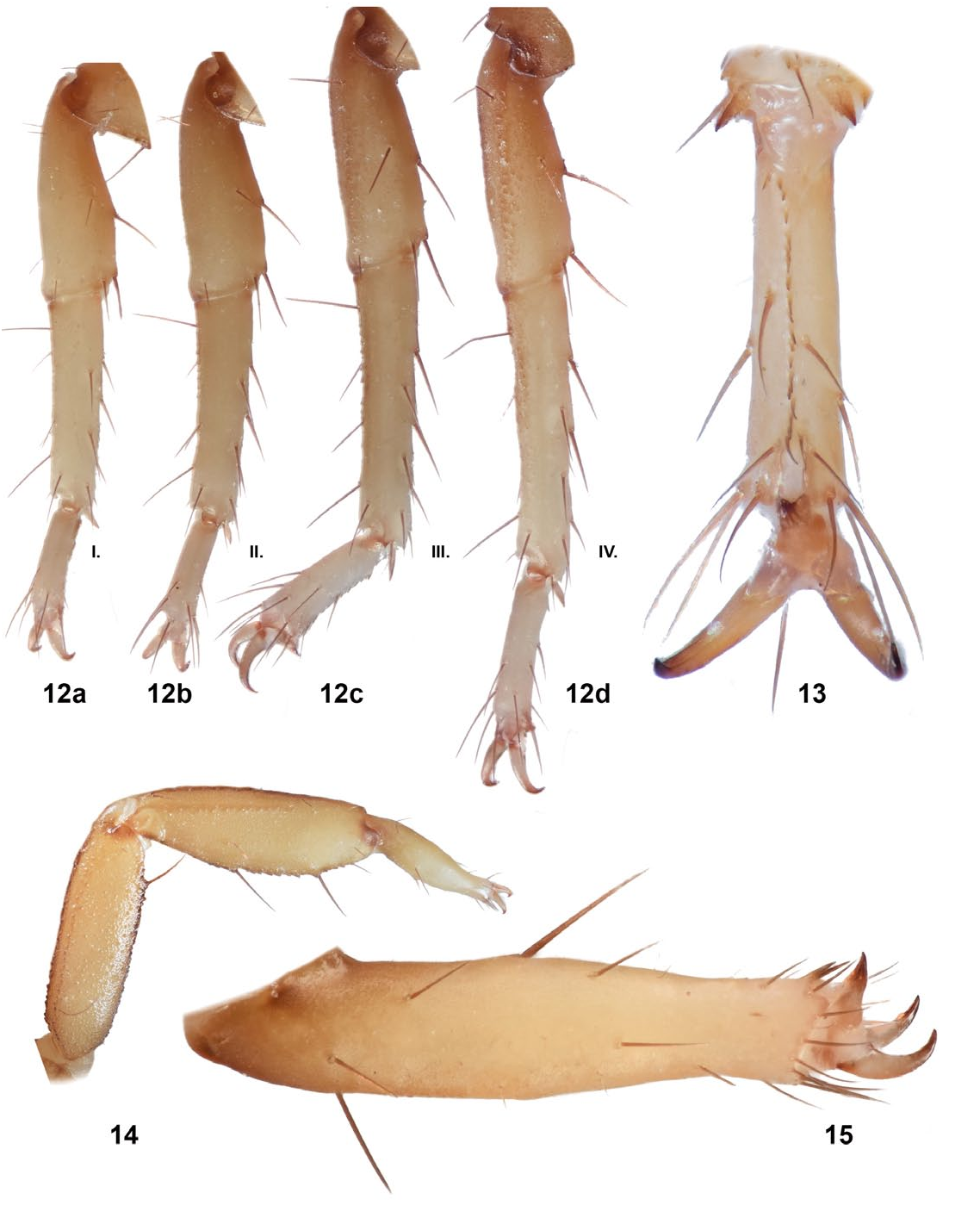
Legs ( Figs. 12–15 View Figures 12–15 ). Ventral surface of tarsomere I with single median row of spinules terminating distally with one spinule pair.An anomaly was discovered on left leg III ( Figs. 14 & 15 View Figures 12–15 ). It appears that the basitarsus and telotarsus never completely developed. These segments are completely missing with the apotele growing directly from the tibia. The tibia appears to be mostly complete, however the patterns of macrosetae are not consistent with the tibia on other legs. Numerous macrosetae are situated at the distal edge of the tibia. While references of limb regeneration in scorpions appear to be rare, Maruzzo & Bortolin (2013), did provide a similar observation, noting that only the apotele appears to regenerate.
Metasoma ( Fig. 11 View Figures 5–11 ). Carapace of male is longer than the fifth metasomal segment. Ratio of segment I length/width 0.80; of segment II length/width 0.81; of segment III length/width 0.88; of segment IV length/width 1.10; of segment V length/ width 1.58. Segments I−IV: dorsolateral carinae strong and granular with distal denticle of I−IV enlarged and spinoid. Lateral supramedian carinae I−IV strong and granular with enlarged spinoid distal denticle. Lateral inframedian carinae moderately granular on posterior 4/5 of segments I−II, posterior ½ of III, and weak to obsolete on IV; individual granules increasing in size distally on segments II−III. Ventrolateral carinae moderately granular on I−III; IV strong, granular. Ventral submedian carinae weak to moderate on I− IV. Dorsal and lateral intercarinal spaces very finely granular. Segment I−IV ventral submedian setae 3:3:3:3. Segment V: Dorsolateral carinae moderate, distally crenulate, basally granular. Lateromedian carinae weak and granular on proximal 3/5, obsolete on distal 2/5. Ventrolateral and ventromedian carinae strong. Intercarinal spaces finely granular. Segment V ventrolateral setae 4/4.
Telson ( Figs. 7–8 View Figures 5–11 ). Rough with 4 pairs of large setae on the ventral surface, 3 large setae along both lateral edges of the vesicle and numerous smaller setae. Prominent, spinoid subaculear tubercle present. Small, triangular granule positioned between subaculear tubercle and base of aculeus. Lateral aculear serrations present, numbering 4 on either side of aculeus.
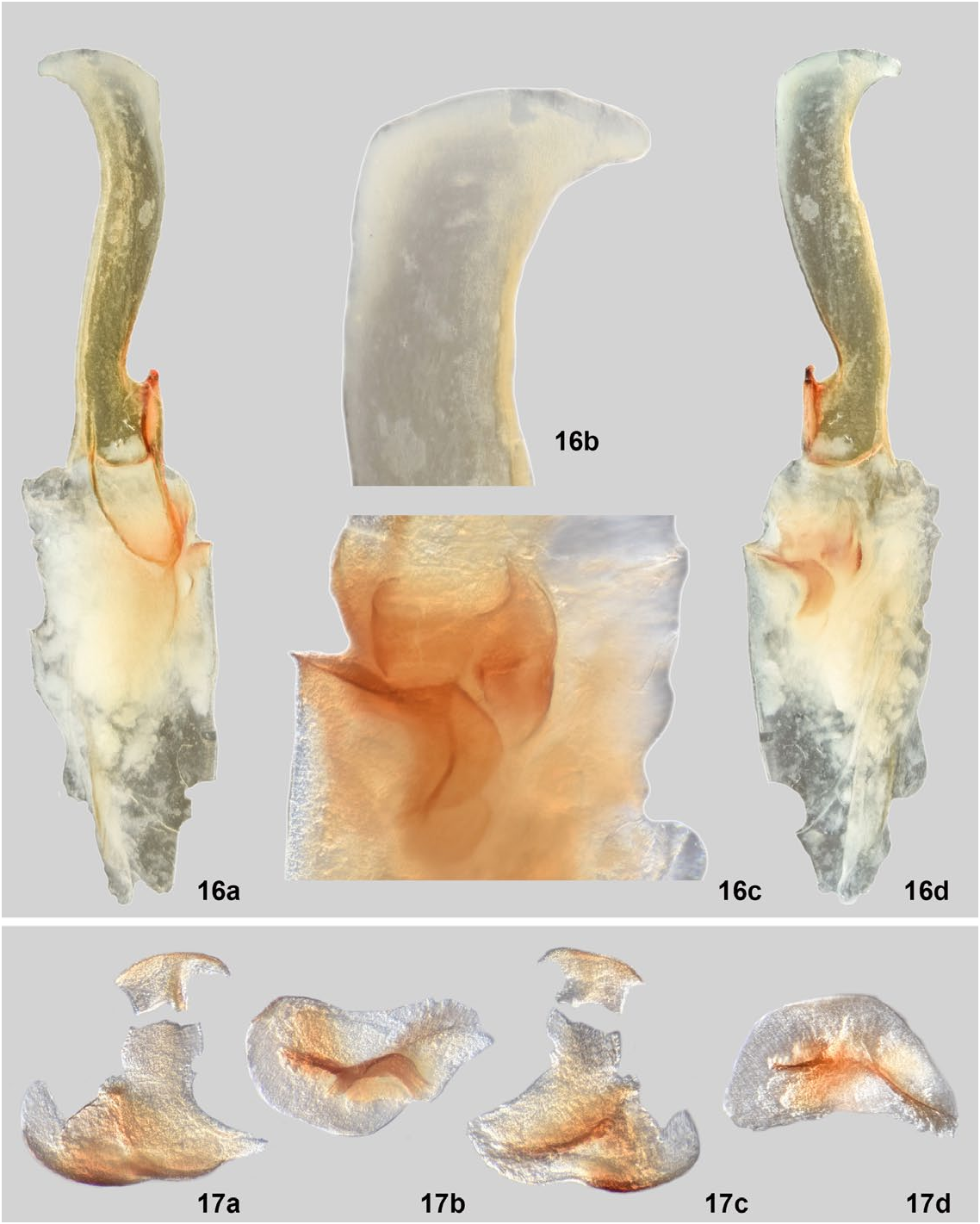
Hemispermatophore ( Fig. 16 View Figures 16–17 ). All descriptions based on right hemispermatophore. Fairly narrow hemispermatophore as shown by trunk and lamina width. Strong basal constriction where lamina widens distally and ends in a wide projection at distal edge. This projection is not seen in the descriptions of either Wernerius inyoensis Webber et al., 2012 , or W. spicatus ( Haradon, 1974) , the only two other known species in the genus. Webber et al. (2012), in their description of W. inyoensis discussed the tapering of the lamina of both W. inyoensis and W. spicatus , showing the ratios of lamina width at distal end of lamina/width at lamina midpoint as 0.900 & 0.652, respectively. In W. mumai , this ratio is 1.871. Lamellar hook sclerotized, weakly bifurcated at distal tip. Shallow dorsal trough.
Measurements (mm): trough difference, 0.46; lamellar hook length, 0.87; lamina length, 2.39; trunk width, 0.60; lamina width, 0.33; ratio of lamellar hook length to lamina length, 0.36; ratio of trough difference to lamellar hook length, 0.53.
Mating Plug ( Fig. 17 View Figures 16–17 ). Sclerotized mating plug with wide base. Stem of average width. Smooth barb, with one tine significantly longer than the other. During dissection, the barbed tip was severed from the stem. In the close-up image of the embedded mating plug, it can be seen that the barbed tip was actually disjointed before the mating plug was removed.
SEXUAL DIMORPHISM is evident, primarily in the body shape, pedipalp ratios, and pectines. The pedipalp fixed finger usually has 6 ID denticles and the movable finger has 7. The mesosoma of the females are wider than those of the males. The pedipalp femur, patella and chela of the male is broader, while female’s pedipalps are more slender. The mean pectinal tooth count for males is 12.31 [n=16], SD = 0.464, for females 11.40 [n=10], SD = 0.516. Also, the pectines of the male are both longer and wider, while females have 1 to 2 of the most proximal teeth smaller and lacking sensorial area, making it easy to distinguish the two sexes.
ECOLOGY.The topotype specimen was found using a blacklight at night at Gold Road, Mohave County, Arizona (35.04119ºN 114.37191 Wº) at an elevation of 986 m a. s. l. The vegetation type is low desert scrub (see Figs. 18–19 View Figures 18–20 ). This species lives under rocks and on canyon walls (see Fig. 19 View Figures 18–20 ). Paravaejovis spinigerus (Wood, 1863) , Stahnkeus subtilimanus (Soleglad, 1972) , and Serradigitus wupatkiensis (Stahnke, 1940) were found syntopically with Wernerius mumai during 12 field trips to Gold Road.
Wernerius mumai exhibit bimodal periods of activity. They are found active on the surface most often in March and April and again in September and October. The authors have not observed surface activity with this species in any other months of the year. This information represents a total of 25 field trips to the topotype locality as well as a second locality discovered by Graeme Lowe while collecting with Wendell Icenogle, along Old Kingman Highway (Black Mountains, SW of Union Pass, 29 August 1992). This bimodal pattern appears to be related to the extreme summer temperature highs found in both localities. According to NOAA (National Oceanic and Atmospheric Administration, USA), the average high air temperatures, per month, at the type locality are: January, 67°F; February, 73°F; March, 83°F; April, 89°F; May, 98°F; June, 108°F; July, 112°F; August, 110°F; September, 103°F, October, 90°F; November, 78°F; and December, 65°F. This species appears to be surface active when daytime highs are in the 80’s. This would be from March until early in May and again from late August through October.
REPRODUCTION. Two adult females were kept alive to determine 1 st instar behavior. Both gave birth with the average number of 1 st instars being 12.00 [n=2]. The 1 st instar orientation on the mother’s back was generally non-random, as is seen with many species of Vaejovis ( Ayrey, 2012, 2013a, 2013b). Most were facing anteriorly with the prosoma down and the metasoma raised over the prosoma of the juvenile immediately posterior. The photo above, taken 24 hours after birth, shows 8 juveniles displaying the standard orientation for the tribe Stahnkeini , while what appears to be the last 5 instars oriented randomly. This type of orientation has been observed, by the authors, in some other genera of Vaejovidae .
DISTRIBUTION. Known only from the Black Mountains , Mohave County, Arizona .





| USNM |
Smithsonian Institution, National Museum of Natural History |
| RFA |
Universidade Federal do Rio de Janeiro |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Wernerius mumai ( Sissom, 1993 )
| Ayrey, Richard F. & Myers, Brandon T. 2020 |
Vaejovis mumai
| SISSOM 2000: 551 |
| SISSOM 1993: 65 |