
Xystosomus gruti Bates, 1871
|
publication ID |
https://doi.org/10.5479/si.00810282.140 |
|
DOI |
https://doi.org/10.5281/zenodo.3848473 |
|
persistent identifier |
https://treatment.plazi.org/id/61278799-FFE4-FF82-DCEC-FE66F97E47AE |
|
treatment provided by |
Plazi |
|
scientific name |
Xystosomus gruti Bates |
| status |
|
FIGURES 3 View FIGURE 3 , 22 View FIGURE 22 a-c, 23 24 View FICURES 23-30 , 36 View FIGURE 36 , 69 View FIGURE 69
Xystosomus grutii Bates, 1871a:248 View in CoL . [Lectotype, female, in MHNP, here selected. Type-locality: Rio de Janeiro, Brazil.]
Xystosomus hilaris Bates, 1871b:266 View in CoL . [Holotype, male, in MHNP ( a specimen mentioned by Bates). Type-locality: Ega (Teffe) , Brazil. New synonymy.]
Xystosomus belti Bates, 1878:601 View in CoL . [Lectotype, male, in MHNP, here selected. Type-locality: Chontales, Nicaragua. New synonymy.]
Xystosomus olivaceus Bates, 1878:601 View in CoL . [Holotype, female, in MHNP, a specimen mentioned by Bates, 1882:147, where he recognized his species as a color aberration of X. belti Bates. View in CoL Type-locality: Chontales, Nicaragua.]
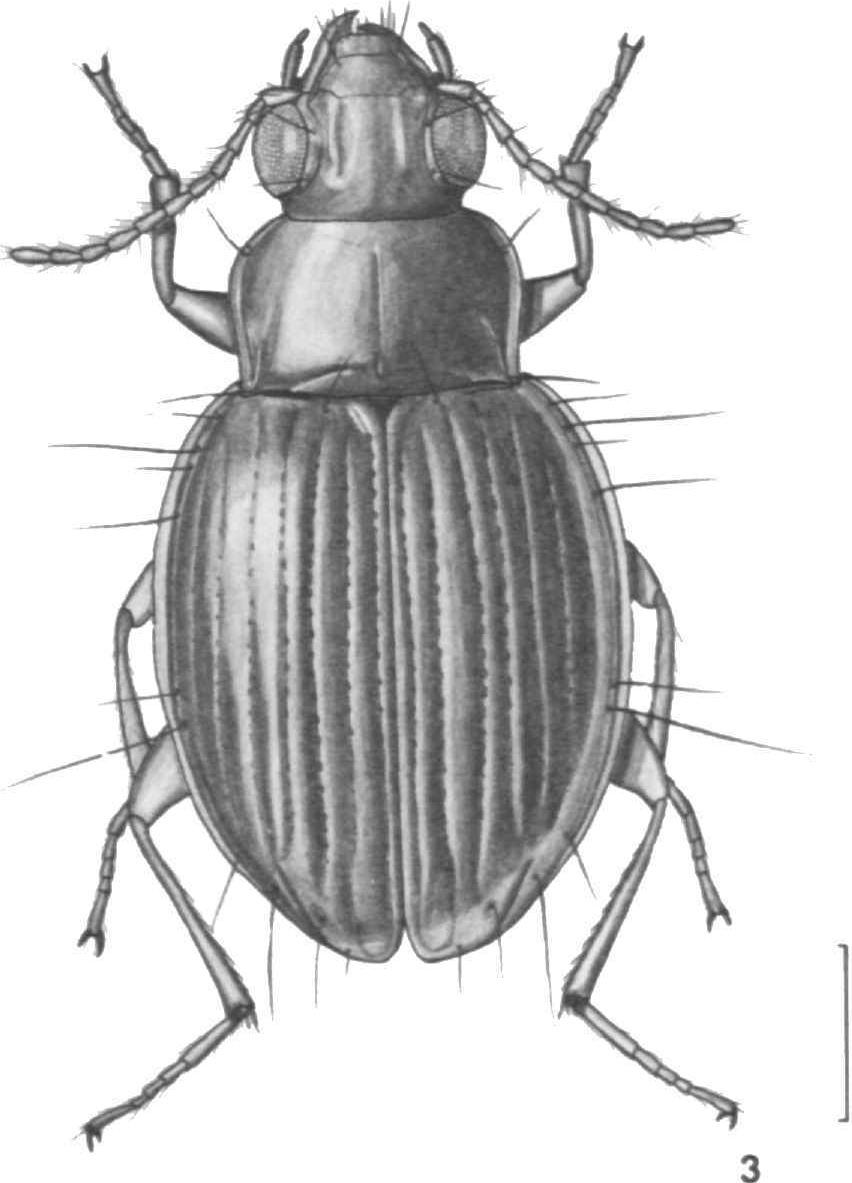
DESCRIPTION.—Form ( Figure 3 View FIGURE 3 ): Large, broad, moderately convex (much less so than members of laevis group) with large head, narrow pronotum (compared with members of laevis group), and fully striate elytra. A variable species (see below) and easily distinguished from the similar X. ampliatus by the deeper lateral striae and the green metallic luster of the entire dorsal surface of the pronotum and elytra.
Color: Head and venter rufopiceous; pronotum and elytra dorsally with metallic green luster; appendages testaceous or partly infuscated.
Head: Broad between eyes; frontal furrows moderately impressed; eyes very large and prominent.
Pronotum ( Figure 3 View FIGURE 3 ): Transverse (W/ L, x 1.60; range, 1.41-1.85; 69 specimens); lateral setigerous pores present just anterior to middle and at hind angles; laterobasal carina well developed; hind angles about 90° and sharp; side margins broadly explanate and more or less straight to apical third.
Elytra: Striae 1-8 of each elytron well impressed and punctulate, sutural stria and lateral four striae more deeply impressed and more coarsely punctulate than striae 2-4; side margins broadly explanate; humeral projection well developed, blunt; chaetotaxy formula Eo-la, 2b, 3b, 4c, 5a, 6a, 7, 8a, and Ed- 1, 7b, 8; plica well developed externally.
Microsculpture: Frons ( Figure 22 a View FIGURE 22 ) with wellimpressed, nearly isodiametric reticulation; pronotum ( Figure 22b View FIGURE 22 ) and elytra ( Figure 22 c View FIGURE 22 ) with fine transverse lines which fork but do not form meshes .
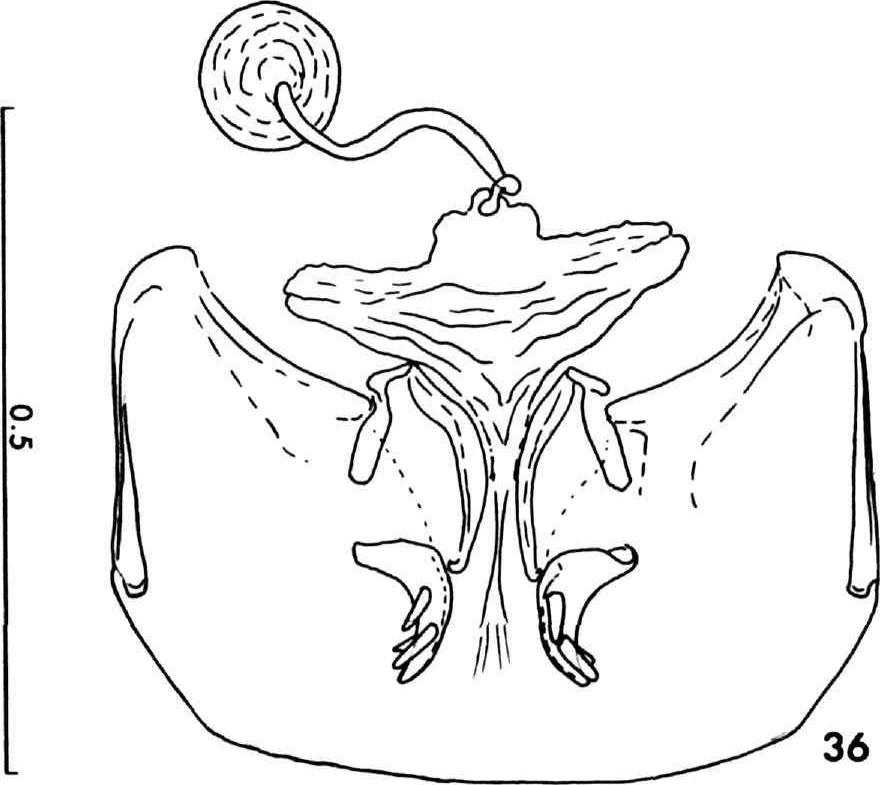
Secondary sexual characters: Male genitalia characteristic of the species group (Figure 23). Female genitalia characteristic of the species group ( Figure 36 View FIGURE 36 ).
Size: Sixty-nine specimens: length, 3.7-5.2 mm; width, 1.9-2.5 mm.
VARIATION.—The depth of the elytral striae varies from deep to shallow. Northern South America (Peru) is a center for specimens with deep striae.
To the north of this center, shallow or deep striae are found on Central American specimens, but only shallow striae are found on the Mexican specimens.
To the south, either shallow or deep striae are found on the Brazilian specimens. Many more samples are needed to accurately assess this characteristic, but from preliminary data it appears that depth of striae is bimodal with shallower striae in the more temperate climates of the species' range.
Slight variation occurs in the shape of the apex of the male median lobe of the genitalia (Figures 23, 24). This variation appears within population samples, however, and is not correlated with geographical area.
The great difference in size (see above) is also independent of geographical area. Both large and small individuals were collected in Costa Rica and Panama. All specimens from Brazil are large, but this may be sample bias, as there are only nine specimens.
NATURAL HISTORY.— In August, in Mexico, George Ball and I collected a male specimen that we saw running on the sun-lit bark of a fallen and partially burnt "buttress tree" after we disturbed some bracket fungi. On Barro Colorado Island, in December, my wife and I collected a female specimen in a pile of deep loose leaves under the crown of a recently fallen tree. The latter beetle was in the company of X. nigripalpis (see below for details). F. Nevermann's excellent collecting records pinned with each specimen give the following data (translated from German): "on leaf pile in sawmill, June," 2 specimens; "on leaves of Cedrela mexicana, May" ( Meliaceae ), 4 specimens; "on wilted foliage of Quararibae turbinata, October" ( Bombacaceae ), 10 specimens; "on dry wood of Pentaclethra filamentosa , March" (Leguminosae), 1 specimen; "on dry wood of Virola war- burgii, February" ( Myristicaceae ), 2 specimens; "under loose bark, November and June," 4 specimens; "wilted leaf of Acanthorhiza sp., June" ( Phoenicaceae ), 1 specimen; "at fermenting plant juice on freshly cut wood, August," 5 specimens; "at light, July," 1 specimen. I have observed a captive female from Barro Colorado Island in flight.
Further records indicate specimens were collected in September in Brazil, but no teneral specimens were seen to indicate at what season immatures might be discovered.
The altitudinal range of X. gruti is from near sea level on Barro Colorado Island to about 4,000 feet (1,219 m) on Volcan de Chiriquf, with most intermediate elevations represented on specimen labels.
In summary, this is a widespread and variable species occurring at low and medium elevations. In habits, it is probably arboreal, or at least subarboreal, and is capable of flight. Adults were collected in every month except January and April. It is probable that adults and immatures overlap.
BEHAVIOR.—The specimen from Barro Colorado Island was collected alive and was returned to Washington for further study (it is still alive at this writing). With only one individual, intraspecific reactions are not possible, but when this specimen and X. nigripalpis specimens are placed together in the same petri dish both demonstrate fierce aggressiveness. This reaction also occurs between members of X. nigripalpis and happens when two beetles come within "setal range" of each other (the elytral "Eo" setae on these beetles are very long). The result of contact is several quick lunges with the mandibles directed toward the other beetle. Many of my living specimens of X. nigripalpis are missing the apical articles of the antennae.
DISTRIBUTION ( Figure 69 View FIGURE 69 ).—The range of this species extends from Vera Cruz, Mexico, to Rio de Janeiro, Brazil. Throughout the range these beetles are distributed both at lower and middle elevations and in lowland tropical forests and cloud forests.
LOCALITY RECORDS ( Figure 69 View FIGURE 69 ).—I have seen at least 74 specimens (old cotypes in BMNH and MHNP not counted) from the following localities:
MEXICO: VERACRUZ: 2.5 miles west of Sontecomapan
( UASM); C 6rdoba (MHNP).
CENTRAL AMERICA: BRITISH HONDURAS: Toledo District ( MCZ). COSTA RICA: Hamburg Farm at Reventaz6 n ( USNM); Iberia Farm in Santa Clara Province ( USNM); Las Mercedes ( USNM). NICARAGUA: Chontales ( MHNP). PANAMA: Barro Colorado Island in Canal Zone ( USNM); Bugaba ( BMNH, MHNP); Cabima ( USNM); Volcan de Chiriqui ( BMNH, MHNP).
SOUTH AMERICA: BRAZIL: Amazonas Province: Teffe ( MHNP); Itaituba ( MHNP). Para Province: Para ( MHNP). Rio de Janeiro Province: Lagoa de Saquarema ( MHNP); " Rio Janeiro " ( MHNP); Rio Parahyba ( MHNP). Santa Catarina Province: Hansa ( MHNP). FRENCH GUIANA: Gourdonville ( MHNP). PERU: Monson Valley at Tingo Maria ( CAS ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Bembidiini |
|
SubTribe |
Tachyina |
|
Genus |
Xystosomus gruti Bates
| Erwin, Terry L. 1973 |
Xystosomus belti
| Bates 1878: 601 |
Xystosomus olivaceus
| Bates 1878: 601 |
Xystosomus grutii
| Bates 1871: 248 |
Xystosomus hilaris
| Bates 1871: 266 |