
Smittina cervicornis ( Pallas, 1766 ), 1968
|
publication ID |
https://doi.org/10.11646/zootaxa.4545.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:4F0A6836-1EC0-4D9F-9963-8C73358BCCBA |
|
DOI |
https://doi.org/10.5281/zenodo.6488945 |
|
persistent identifier |
https://treatment.plazi.org/id/5F6687C4-1D21-5302-FF34-FF7A4717379B |
|
treatment provided by |
Plazi |
|
scientific name |
Smittina cervicornis ( Pallas, 1766 ) |
| status |
|
Smittina cervicornis ( Pallas, 1766)
( Figs 1 View FIGURES 1–3 , 4–22 View FIGURES 4–9 View FIGURES 10–16 View FIGURES 17–22 ; Table 2)
Millepora cervicornis Pallas, 1766: 252 .
Smittia colletti Jullien in Jullien & Calvet, 1903: 100 , 149, pl. 12, fig. 3.
Smittia cervicornis (Ellis & Solander) : Jullien & Calvet, 1903: 151, pl. 17, fig. 6a–d.
Smittia cervicornis (Pallas) : Barroso, 1912: 42, fig. 8.
Porella cervicornis (Pallas) : Calvet, 1931: 93; Gautier, 1962: 204.
Smittina cervicornis (Pallas) : Cook, 1968: 210; Zabala & Maluquer, 1988: 121, fig. 266, pl. 12C; Hayward & McKinney, 2002: 49, fig. 22A–C.
Smittina colletti (Jullien & Calvet) : Reverter-Gil & Fernández-Pulpeiro, 2001: 109.
?Not Porella colletti (Jullien & Calvet) : Gautier, 1962: 206; Harmelin, 1969: 297, fig. 5:4.
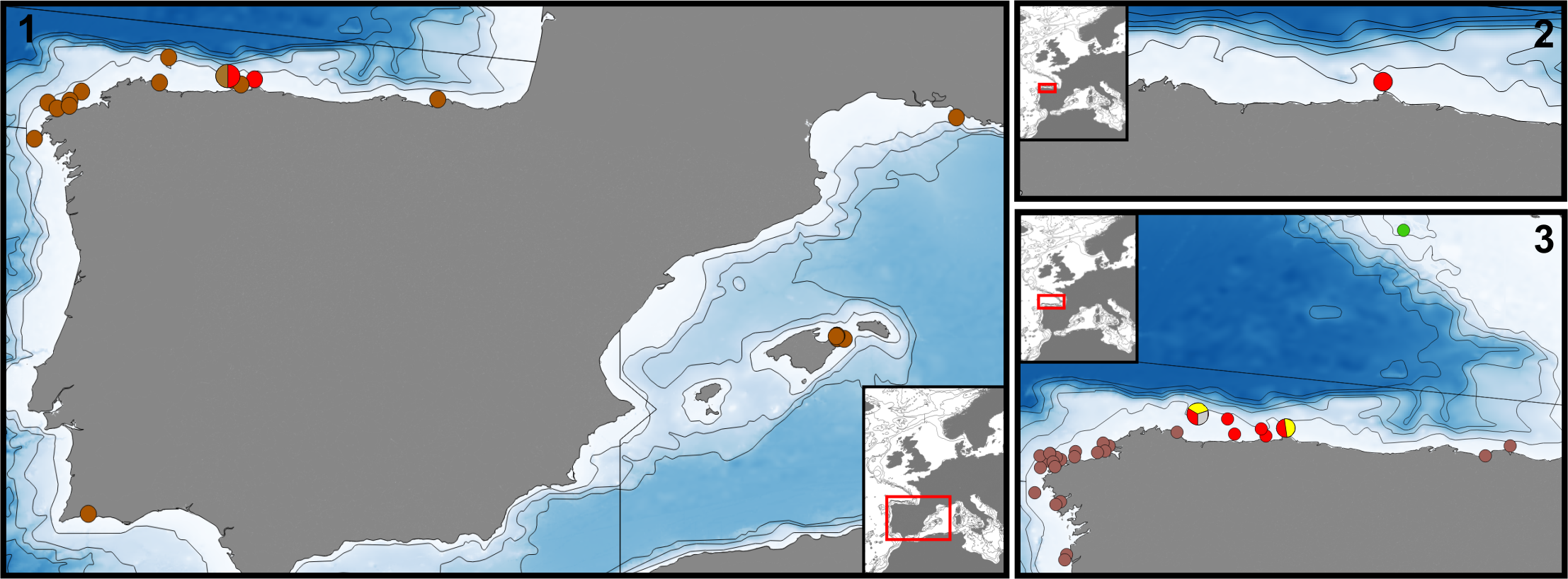
Material examined. Holotype (by monotypy) of Smittia colletti : MNHN 3913 About MNHN , Hirondelle st. 57, 43.74167°N, 06.20444°W, 240 m depth, Bay of Biscay, Calvet Coll. GoogleMaps
Additional material: MOM 420122 View Materials , Hirondelle st. 53, 43.74722°N, 05.86278°W, 135 m depth, Bay of Biscay, Calvet Coll. as Smittia colletti GoogleMaps . MOM 420180 View Materials : Hirondelle st. 57, 43.74167°N, 06.20444°W, 240 m depth, Bay of Biscay, Calvet Coll. as Smittia colletti GoogleMaps . MOM 420183 View Materials : Hirondelle st. 57, 43.74167°N, 06.20444°W, 240 m GoogleMaps depth, Bay of Biscay , Calvet Coll. as Smittia cervicornis . MHNUSC-Bry 422: Point Etxandarri, Bay of Biscay, 43.44225°N, 02.92000°W, 25 m GoogleMaps depth. MHNUSC-Bry 423: Avilés Canyon , Bay of Biscay, 43.6709°N, 06.07703°W, 86 m GoogleMaps depth. MHNUSC-Bry 534: one colony, A Coruña , 43.56056°N, 08.63361°W, 147 m depth, 12/ 04/2003 GoogleMaps . MHNUSC-Bry 536: fragments, Malpica , 43.42528°N, 08.82361°W, 119 m depth, 16/11/2001 GoogleMaps . MHNUSC-Bry 535: two small colonies, Malpica , 43.39667°N, 09.18139°W, 186 m depth, 02/11/2003 GoogleMaps . MHNUSC- Bry 537: fragments of colonies, Corme, 43.31083°N, 09.03083°W, 100 m depth, 31/12/2002 GoogleMaps . MHNUSC-Bry 538: a small colony, Fisterra , 42.84778°N, 09.39750°W, 152 m depth, 08/12/2002 GoogleMaps . MHNUSC-Bry 426: Portimão , 37.13611°N, 08.52722°W, 27/03/2004 GoogleMaps , many specimens in fishing nets. MNCN 25.03 About MNCN /4142: Portimão , 37.13611°N, 08.52722°W, 27/03/2004 GoogleMaps , many specimens in fishing nets. MHNUSC-Bry 424: Menorca Channel , st. P52, 39.85008°N, 03.48209°E, 69 m depth, 12/09/2011 GoogleMaps . Western Mediterranean ; MHNUSC-Bry 449: Imperial de Terre, Riou Island, 43.1729°N, 5.39327°E, 20–22 m depth, 28/09/2018, coll. A. Ostrovsky. GoogleMaps MHNUSC-Bry 450: Imperial de Terre , Riou Island, 43.1729°N, 5.39327°E, 20–22 m depth, 28/09/2018 coll. A. Ostrovsky. GoogleMaps
Additional localities studied (specimens not stored): O Barqueiro, 43.69806°N, 07.39083°W, 68 m depth, 13/ 12/2002. Malpica , 43.35222°N, 08.83778°W, 23 m depth, 02/06/2002 GoogleMaps . Menorca Channel , st. P5, 39.80500°N, 03.49200°E, 56 m depth, 02/09/2011 GoogleMaps . Menorca Channel , st. P6, 39.80620°N, 03.49070°E, 55 m depth, 02/09/2011 GoogleMaps . Menorca Channel , st. P14, 39.7942°N, 03.60070°E, 72 m depth, 04/09/2011 GoogleMaps . Menorca Channel , st. P50, 39.8507°N, 03.47450°E, 68 m depth, 12/09/2011 GoogleMaps . Menorca Channel , st. P51, 39.8507°N, 03.48000°E, 69 m depth, 12/09/2011 GoogleMaps . Menorca Channel , st. P54, 39.83275°N, 03.47425°E, 60 m depth, 13/09/2011 GoogleMaps .
Description. Colony erect, rigid, up to 10–15 cm in height, orange ( Figs 4–5 View FIGURES 4–9 ). Colonies with abundant branches, branching pattern by bifurcation. Branches bilaminar, flat-sectioned, with up to 12 lineal series of zooids by layer, widest just before bifurcations ( Figs 9 View FIGURES 4–9 , 22 View FIGURES 17–22 ). In old parts of the colony frontal budding occurs after secondary calcification of the first layer. Frontal budding gave successive layers with the zooids in irregular disposition ( Figs 21, 22 View FIGURES 17–22 ). Autozooids quadrangular, hexagonal or irregular in shape in successive frontal layers; separated by sutures between slightly raised thin ridges; frontal shield umbonuloid, flat or lightly convex ( Figs 6– 16 View FIGURES 4–9 View FIGURES 10–16 ). Large frontal pores along the frontal surface, being larger in the central area ( Figs 9–17 View FIGURES 4–9 View FIGURES 10–16 View FIGURES 17–22 ). Primary orifice semicircular with a short, broad lyrula in the proximal border ( Figs 17–20 View FIGURES 17–22 ). No spines. Secondary orifice very variable, depending on the degree of secondary calcification and frontal budding. Initially the orifice is keyholeshaped, with a larger distal section corresponding to the primary orifice, and a smaller proximal area, where the suboral avicularium is supported ( Figs 10, 11 View FIGURES 10–16 , 18, 20 View FIGURES 17–22 ). As the calcification increases, the secondary orifice grows as a peristome, initially with a proximal channel toward the avicularium but quickly forming a thickly calcified peristome, cylindrical, projecting from colony surface. In heavily calcified colonies, only a perfectly round orifice persists, with circular growing marks ( Figs 8 View FIGURES 4–9 , 13 View FIGURES 10–16 , 21 View FIGURES 17–22 ).
Avicularium suboral, prominent, with rounded rostrum and a semicircular mandible, directed proximally. Distal part of avicularium with a raised cup. Crossbar complete, with a robust columella which is frequently bifurcate ( Figs 18–20 View FIGURES 17–22 ). Ovicell not seen in studied specimens.
Remarks. Jullien (in Jullien & Calvet 1903) described Smittia colletti based on one colony from st. 53 of the l’Hirondelle (Bay of Biscay, 135 m depth). This sample is the holotype of the species and is now stored at the MNHN. Later, Calvet (in Jullien & Calvet 1903) reported two isolated fragments from st. 57 ( 240 m depth). Smittia colletti was never recorded again in the Atlantic, but the original records were compiled by Reverter-Gil & Fernández-Pulpeiro (2001) as Smittina colletti . In the Mediterranean, one specimen was reported by Gautier (1962) at 162 m depth in the Banc du Esquerquis ( Tunisia), and Harmelin (1968, 1969) recorded specimens from two other Mediterranean localities: Strait of Sacarpanto, 29–33 m depth and 60–80 m depth, and in Santorini at 100– 110 m depth. Other records were included as nominal references by Harmelin (1979), Zabala & Maluquer (1988), Rosso (2003) and Rosso & Di Martino (2016), but apparently no more material was studied.
Calvet (in Jullien & Calvet 1903) also recorded four samples of S. cervicornis at the locality where he recorded S. colletti (st. 57). Smittina cervicornis is a common and abundant species in the Mediterranean, where it was described by Pallas (1766) and later recorded by several authors (e.g. Calvet 1907, 1927, 1931; Gautier 1962; Zabala & Maluquer 1988; Álvarez 1994; Harmelin et al. 1994; Hayward & McKinney 2002; Rosso 2003; Sanfilippo et al. 2013; Rosso & Di Martino 2016), but the type material of the species, described from Riou Island (Marseille, France) appears to be lost, as is the rest of the Pallas collection ( Kukliński & Taylor 2008). Outside the Mediterranean, S. cervicornis was also recorded from different localities: Morocco ( Canu & Bassler 1925, 1928; d’Hondt 1978), Gulf of Cádiz (López de la Cuadra 1991; Álvarez 1994), south of Portugal ( Reverter-Gil et al. 2014), Canary Islands ( Arístegui Ruíz & Fernández Gil 2003) and tropical west Africa ( Cook 1968), and also from the Bay of Biscay ( Jullien & Calvet 1903; Calvet 1907; Barroso 1912; Aguirrezabalaga et al. 1986; Álvarez 1991).
Although S. cervicornis and S. colletti , collected at the same locality, seemed very similar, Jullien & Calvet (1903) did not discuss the differences between the two species. It is a mystery why, of all the material collected at st. 57, Calvet (in Jullien & Calvet 1903) decided to report four of the samples as S. cervicornis , whereas he preferred to identify two fragments as a different species, S. colletti . Perhaps he was simply retaining an earlier, unpublished identification of the sample made by Jullien, who was actually the person who described the species in the first part of the work. The first author to provide diagnostic characters for S. colletti was Gautier (1962), who stated that the species comprises white colonies with fewer and narrower branches, and with smaller zooids, and that it seems to live in deeper waters than S. cervicornis . However, these statements were based on material collected in the Mediterranean and were not reported in the original description of S. colletti . More importantly, these features are not visible in the original material of the species.
We have studied the original specimens of S. colletti ( Figs 6, 7, 9–11 View FIGURES 4–9 View FIGURES 10–16 , 18, 20 View FIGURES 17–22 ), together with other newly collected material from several localities in the Bay of Biscay between 25 and 86 m depth, off the NW Iberian Peninsula between 23 and 186 m depth, in the Balearics between 55 and 72 m depth and specimens collected at 20– 22 m depth from Riou Island, Marseille ( type locality of S. cervicornis ); and also several colonies collected in the Algarve (south Portugal) previously reported by Souto et al. (2010). Our observations were compared with existing descriptions of S. cervicornis (e.g. Gautier 1962; Zabala 1986; Zabala & Maluquer 1988; Hayward & McKinney 2002). All the material shows wide inter- and intra-colony variability. This pertains especially to frontal-shield morphology, mainly correlated with secondary calcification. It also involves the peristomial area, which is initially keyhole-shaped with a variable coarse calcification that changes to form a circular rim in older zooids. The suboral avicularium seems to be more stable morphologically, but varies in the size of the proximal opesia. There is a welldeveloped central columella, which is typically bifurcate. None of the variations shows a consistent pattern, however, so it is not possible to separate the studied material into different groups. Even the biometrics of all the characters in the colonies studied are similar (see Table 2).
Recently, a new species of Smittina was described from Mauritania, Smittina imragueni Matsuyama et al., 2015 , which closely resembles S. cervicornis . According to Matsuyama et al. (2015) this species differs from S. cervicornis mainly in the position and size of the avicularia, which are smaller and almost perpendicular to the growing plane. Moreover, the frontal shield of the zooids figured by Matsuyama et al. (2015), seem to be more arcuate than in the specimens studied during this work, although this character could be variable and dependent on secondary calcification. Similar differences were highlighted by Matsuyama et al. (2015) to differentiate S. colletti from S. imragueni , but the authors also indicated the presence of one medial denticle on top of the lyrula of S. colletti as a difference. We were unable to observe this denticle in type specimens of S. colletti or in any other specimen studied here, so in our opinion this difference is incorrect.
9909090909090SD, standard deviation; N, number of measurements.
In the absence of significant differences we conclude that the type specimens of S. colletti and the other specimens studied here are actually referable to S. cervicornis , which becomes a senior subjective synonym. Mediterranean material reported by Gautier (1962) and Harmelin (1968, 1969) as Porella colletti probably corresponds to a different species, but confirmation is beyond the scope of the present paper.
Smittina cervicornis seems abundant locally in the Bay of Biscay, forming groups of colonies that were observed between 20 and 30 m depth off Point Etxandarri. It is also very abundant in the Mediterranean, where the species has been described as an important component of coralligenous biocoenoses (e.g. Chimenz & Faraglia 1995; Ballesteros 2006; Harmelin 2017).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Smittina cervicornis ( Pallas, 1766 )
| Souto, J. & Reverter-Gil, O. 2019 |
Smittina colletti
| Reverter-Gil, O. & Fernandez-Pulpeiro, E. 2001: 109 |
Smittina cervicornis (Pallas)
| Hayward, P. J. & McKinney, F. K. 2002: 49 |
| Zabala, M. & Maluquer, P. 1988: 121 |
| Cook, P. L. 1968: 210 |
Porella cervicornis
| Gautier, Y. - V. 1962: 204 |
| Calvet, L. 1931: 93 |
Smittia cervicornis
| Barroso, M. G. 1912: 42 |
Smittia colletti Jullien in Jullien & Calvet, 1903 : 100
| Jullien, J. & Calvet, L. 1903: 100 |
Smittia cervicornis
| Jullien, J. & Calvet, L. 1903: 151 |
Millepora cervicornis
| Pallas, P. S. 1766: 252 |