
Rhinolekos garavelloi Martins & Langeani, 2011
|
publication ID |
https://doi.org/ 10.1590/S1679-62252011000100005 |
|
persistent identifier |
https://treatment.plazi.org/id/5F4887C1-FFC2-CB42-149E-5045FEF531C6 |
|
treatment provided by |
Carolina |
|
scientific name |
Rhinolekos garavelloi Martins & Langeani |
| status |
sp. nov. |
Rhinolekos garavelloi Martins & Langeani View in CoL , new species
Fig. 5 View Fig
Holotype. DZSJRP 10479 , 31.4 mm SL, male, Brazil, Goiás State, Caldas Novas, rio Paranaíba drainage, stream at Fazenda Lageado , near GO-213 road, after rio Corumbá bridge, 17º44’20”S 48º28’27”W, 8 Jun 2007, F. Langeani, L. G. G. Silveira, D. Simiele, M. M. F. Marinho & J. P. Serra. GoogleMaps
Paratypes. All from same locality of holotype. DZSJRP 10477 , 22 , 3 c&s, 15.2-36.2 mm SL (11, 31.2-36.2 mm SL), MCP 44057, 6 View Materials , 24.8-32.2 mm SL (3, 30.6-32.2 mm SL), 11 May 2007, L. G. G. Silveira & F. Langeani. DZSJRP 12191 , 37 , 4 c&s, 10.7-32.8 mm SL (11, 29.8-32.8 mm SL), MNRJ 34127 View Materials , 5 View Materials , 21.4-32.5 mm SL (2, 29.0- 32.5 mm SL), MZUSP 103697 View Materials , 5 View Materials , 20.0- 31.3 mm SL (2, 31.2- 31.3 mm SL), collected with the holotype .
Diagnosis. Rhinolekos garavelloi differs from its congeners by the following characters: shorter postanal length (29.0- 34.6% in SL vs. 34.7-37.7% in R. britskii and 35.7-41.3% in R. schaeferi ); longer thoracic length (18.1-20.8% in SL vs. 14.7- 18.0% in R. britskii and 13.7-17.6% in R. schaeferi ); 30-35 dorsal plates (vs. 24-28 in R. britskii and 26-28 in R. schaeferi ). Furthermore, it could be distinguished from R. britskii by the absence of transverse dark bands in pectoral-, pelvic- and anal-fin rays (vs. presence); by having 32 vertebrae (vs. 31), and anterior portion of the compound supraneural-first dorsal-fin proximal radial contacting the neural spine of the 10 th vertebra (vs. 9 th). It further differs from R. schaeferi by having 23-27 mid-dorsal plates (vs. 18-20); 24-28 mid-ventral plates (vs. 20-22); higher caudal peduncle (10.3-13.0% in SL vs. 7.7-8.7); wider head (24.6-28.1% in SL vs. 20.5-23.9%); premaxillary and dentary accessory teeth absent (vs. present).
Description. Morphometric and meristic data are given in Tables 3 and 4. Dorsal body profile convex from tip of snout to dorsal-fin origin; concave at dorsal-fin base; straight to caudal-fin origin. Small elevation at supraoccipital immediately posterior to eyes. Ventral head profile slightly concave. Ventral body profile almost straight from pectoral-fin origin to analfin origin, ascending at anal-fin base. Greatest body depth at dorsal-fin origin. Greatest body width at opercular region, gradually tapering toward snout and caudal-fin. Head without longitudinal crests, anterior margin rounded in dorsal view. Snout without rostral plate, bearing numerous small plates with thin odontodes; most anterior portion of head naked. Odontodes equal in size and uniformly distributed, not forming rows, on head and body. Eye small; dorsolaterally placed, not visible in ventral view. Iris operculum present. Infraorbital canal entering infraorbital series via compound pterotic. Compound pterotic roughly quadrangular in shape, without elongate posterior extension, its posteroventral margin with irregular and median to large perforations. Supraoccipital not contributing to the dorsal portion of the swimbladder capsule.
Body entirely covered with bony plates, except on ventral part of head, region overlying opening of swim bladder capsule and around anus and pelvic-fin origin.Abdomen covered with small plates irregularly arranged.
Lips roundish, papillose; lower lip larger than upper lip, with papillae gradually smaller towards edges. Maxillary barbel free from lower lip and reduced. Teeth slender, bifid; median cusp larger and rounded, lateral cusp smaller and pointed. Premaxillary teeth 20-34. Dentary teeth 18-31. Number of teeth increasing with size of specimen. Premaxillary and dentary accessory teeth absent.
Dorsal-fin rays ii,6-7; originating at vertical through middle of pelvic fin; its length surpassing anal-fin origin; spinelet small, roughly triangular shaped; locking mechanism non-functional. Anterior portion of compound supraneuralfirst dorsal-fin proximal radial contacting neural spine of 10 th vertebra ( Fig. 1b View Fig ). Pectoral-fin rays i,6, originating immediately posterior to opercular opening, surpassing pelvic-fin origin. Cleithrum and coracoid exposed and supporting odontodes only laterally, near pectoral-fin insertion. Arrector fossae partially enclosed by ventral lamina of coracoid; opening relatively large, extending laterally halfway towards pectoral-fin base. Pectoral-fin axillary slit present only in juvenile specimens. Pelvic-fin rays i,4-5, reaching anal-fin origin when depressed. Pectoral- and given for holotype (H; DZSJRP 10479 ) and range of 30 specimens
(DZSJRP 10477, 12191; MCP 44057; MNRJ 34127; MZUSP
103697); range includes holotype. SD = Standard deviation.
pelvic-fin unbranched rays smaller than branched rays; enlarged odontodes at tip of pectoral-fin unbranched ray and at mesial margin of pelvic-fin unbranched ray. Anal-fin rays i,3-5. Caudal-fin rays i,14-15,i; lower lobe slightly longer than upper lobe, in some specimens; four or five dorsal and 3-6 ventral procurrent rays. Adipose fin and azygous plates absent. Median lateral plates 28-32. Median-plate series complete, from compound pterotic to caudal-fin base, absence of canal in one or two plates near region of tenth median plate in some specimens. Vertebrae 32.
Color in alcohol. Ground color of dorsal surface light brown; posterior portion of head with transverse dark bar. Trunk with four conspicuous bars: first at dorsal-fin origin; second at end of dorsal-fin base; third at vertical through middle anal fin; last at caudal-fin insertion; less conspicuous in adult specimens. Head darker than remaining portion of dorsum. Clear area between tip of snout and nares. Opercle region unpigmented. Lateral portion of body light brown with dark longitudinal stripe, from compound pterotic to caudal-fin origin. Ventral surface of body mostly unpigmented. Dorsal, anal-, pectoral- and pelvic-fins membranes hyaline. Dorsal-fin rays with higher concentration of chromatophores, forming transverse bands. Caudal fin homogeneously dark; tip of lobes unpigmented. Procurrent rays with yellowish white coloration sometimes, extending laterally to caudal peduncle.
Sexual dimorphism. Males with conspicuous urogenital papillae immediately posterior to anus and first pelvic-fin ray with a dorsal expanded flap of skin.
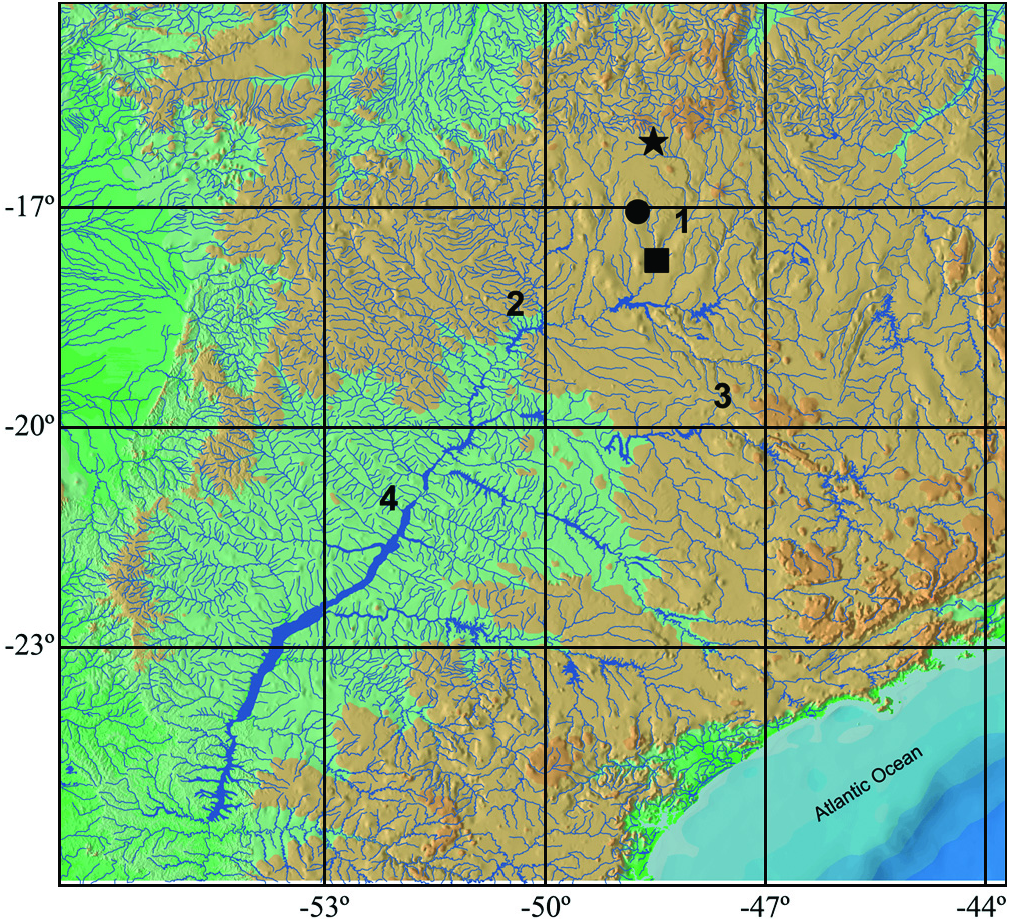
Distribution. Known from the type locality, stream tributary to rio Paranaíba drainage, upper rio Paraná system, Caldas Novas, Goiás State, Brazil ( Fig. 3 View Fig ).
Etymology. Named after Julio C. Garavello, Universidade Federal de São Carlos, for his contributions to the study of Hypoptopomatinae catfishes and to Neotropical Ichthyology.


| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |