
Bactroceras boliviensis Aubrechtová, 2015
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.783.1601 |
|
publication LSID |
lsid:zoobank.org:pub:071EAD63-05ED-4D6C-AC45-8719E6D79E0B |
|
DOI |
https://doi.org/10.5281/zenodo.5795801 |
|
persistent identifier |
https://treatment.plazi.org/id/5F4487AC-FF8F-FF87-FDCD-7BB9FCF378C8 |
|
treatment provided by |
Felipe |
|
scientific name |
Bactroceras boliviensis Aubrechtová, 2015 |
| status |
|
Bactroceras boliviensis Aubrechtová, 2015
Figs 12E, H, J View Fig , 13C View Fig , 24–27 View Fig View Fig View Fig View Fig , 28A–B View Fig , 29B View Fig
Bactroceras boliviensis Aubrechtová, 2015: 206–207 , fig. 6a–d, g.
Bactroceras aff. avus Holm – Marek et al. 2000: 56.
Bactroceras angustisiphonatum (Rüdiger) – Evans 2005: 27–31, pl. 3 figs 12–13, 15, 17–18, pl. 4 figs 2, 16.
Diagnosis
Bactroceras with a large shell diameter (> 45 mm), ornamented with transverse striae, and with concavity of septa of 30%; siphuncle tubular, marginal; diameter of siphuncle ⅛ to ¹/7 of the corresponding conch diameter; connecting ring very thin (after Aubrechtová 2015).
Material examined
Sixty-eight specimens (see Supp. file 1 for list of specimens); one from bed PO 7, two from bed PO 7.5, two from bed PO 123.3, and 62 from bed PO 131, 4 m, 4.5 m, 120.3 m, and 128 m above base of Olenidsletta Member, V1–V2 trilobite zone, Blackhillsian, Floian. An additional number of 15 microscopic specimens from bed PO 131.
Description
The conchs are straight and expand gradually with a low angle of expansion, ca 4° (mean 3.7°; 1 st –3 rd quantile: 2.7–4.5; n = 37) ( Fig. 24 View Fig ). The conch cross section is invariantly circular. The largest specimen in the sample is a fragmentary body chamber with a maximum diameter of ca 48 mm and a length of
92 mm (specimen FMNH-P30179). At the base of the body chamber, where the diameter is 38 mm, a narrow paired dorsomyarian muscle scar is preserved ( Fig. 25A View Fig ).
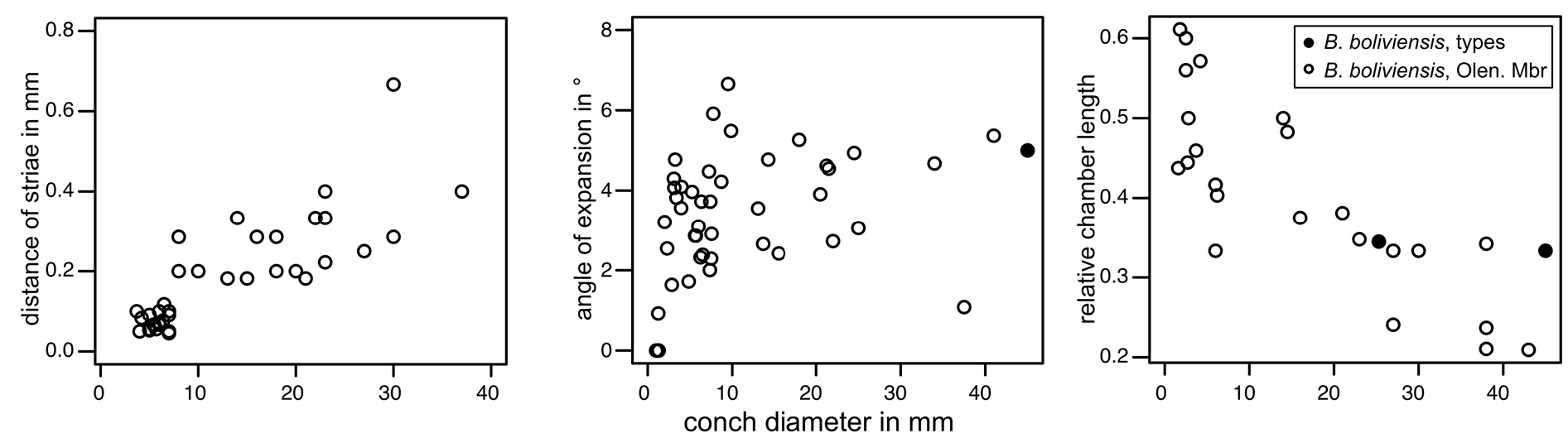
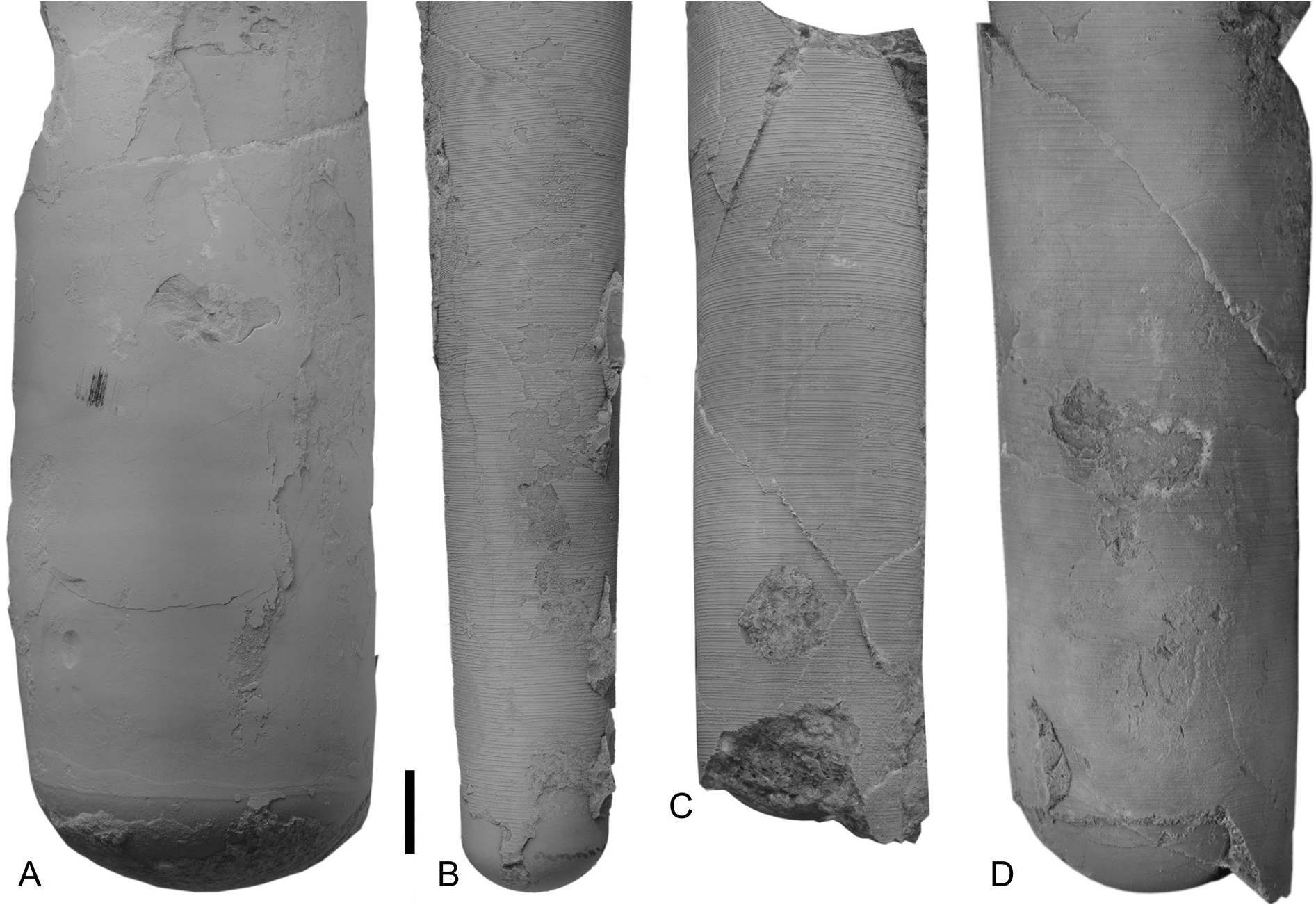
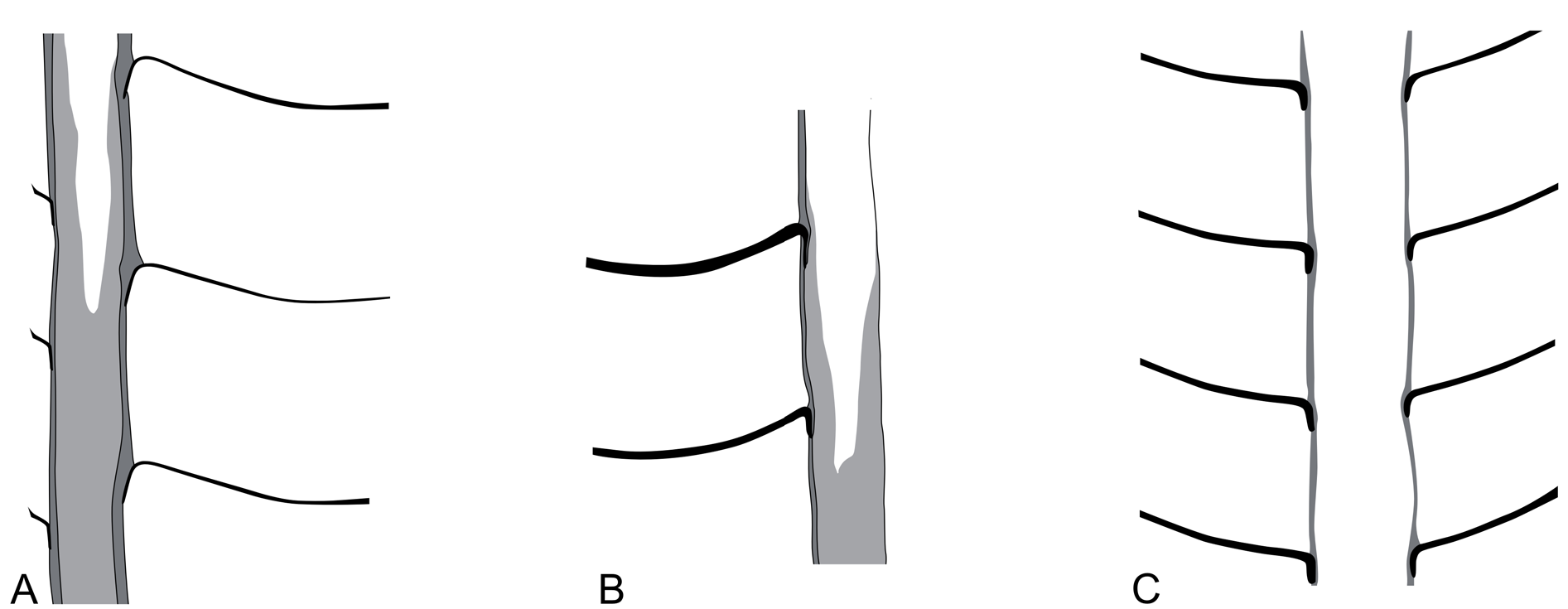
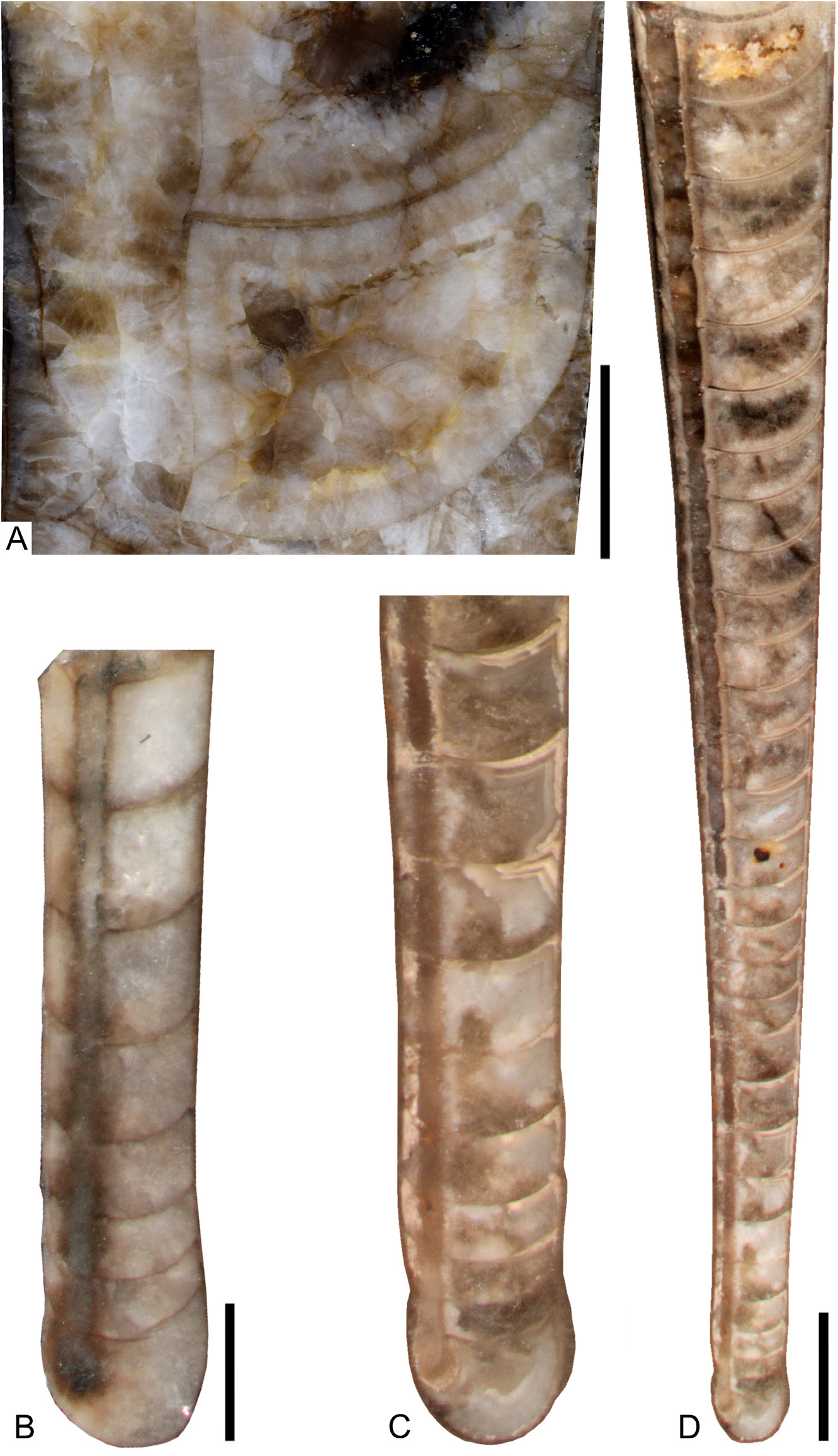
The conch surface is ornamented with striae, which are directly transverse and run parallel to the sutures ( Figs 13C View Fig , 25B–D View Fig ). The striae are very slightly shifted toward the apex at the prosiphuncular side, forming a very shallow ventral sinus. The striae have partly an imbricated profile in well preserved specimens and are rounded in less well preserved and smaller specimens. The spacing of the striae increases with growth in relation to the conch diameter ( Fig. 24 View Fig ), reaching densities between 1–20 lirae per one millimeter.
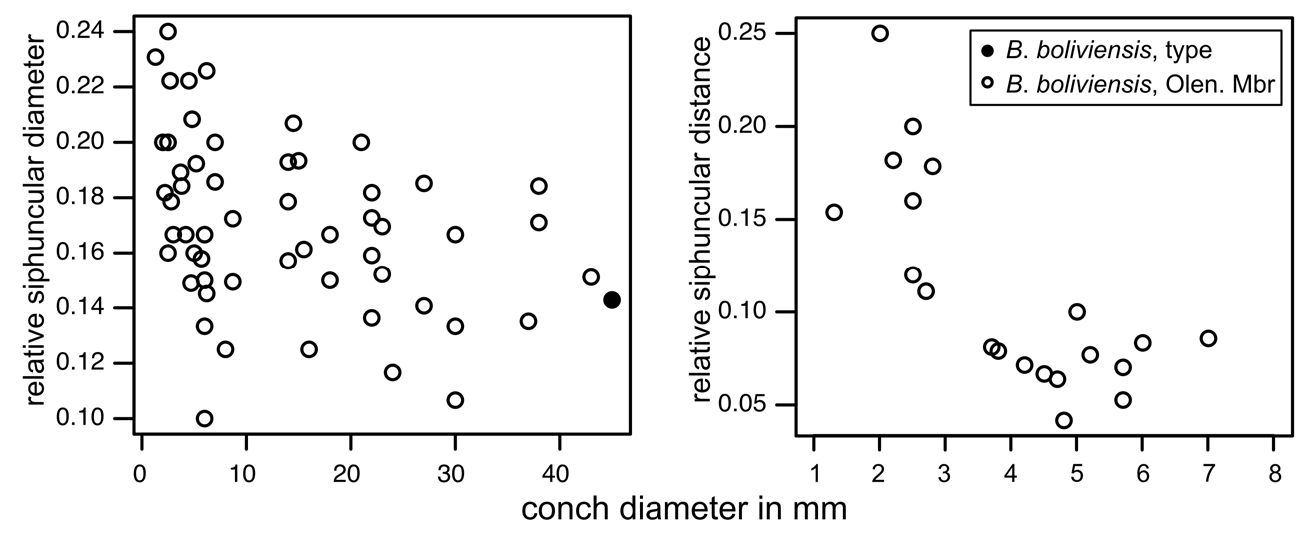
The relative chamber length varies between 0.6 in the smallest preserved growth stages with diameters of less than 5 mm and 0.2 in the largest specimens with diameters> 35 mm. The diameter of the septal foramen is ca ¹/6 of corresponding conch cross section (mean rSD = 0.17; 1 st –3 rd quantile: 0.15–0.19; n = 55) ( Fig. 26 View Fig ). The rSD decreases with conch diameter toward 0.15 at a maximum conch diameter of 43 mm ( Fig. 26 View Fig ). The siphuncle is eccentric, positioned between the center of the conch and the conch margin in early growth stages with conch cross section diameters <6 mm, and it is nearly marginal in later growth stages ( Fig. 26 View Fig ). The septal necks are orthochoanitic and the siphuncular segments are tubular and have a thin connecting ring ( Fig. 27 View Fig ).
The siphuncles of specimen FMNH-P30215 from bed PO 131, and specimens FMNH-P30243 and FMNH-P30259 from bed PO 7.5 ( Figs 27A–D View Fig , 28A–B View Fig ), contain an irregular conical continuous endosiphuncular lining, which adapically fills the entire siphuncle but thins out adorally within a length of ca 20–30 mm at conch diameters of up to 38 mm.
The conch apex consists of a nearly spherical protoconch with a mean diameter of 1.1 mm (1 st –3 rd quantile: 1.1–1.2; n = 15) and a mean length of 1 mm (1 st –3 rd quantile: 1.0–1.1; n = 15), which is distinguished from a shaft by a shallow, often very indistinct constriction ( Figs 12J View Fig , 29B View Fig ). The spherical initial part is smooth, without a cicatrix ( Fig. 12H View Fig ), and the following ca 5–10 mm of the initial part of the conch is nearly smooth. In some well-preserved specimens a faint, narrow, transverse ornamentation is visible, which becomes more pronounced during growth. The shaft and the following ca 5–10 mm are almost straight and have a very low angle of expansion or are tubular. At a distance of 25 mm from the apical tip the conch is only 2.7 mm wide in specimen FMNH-P30165. The caecum is elongated; in specimen FMNH-P30176 it is ca 0.4 mm long and 0.3 mm wide, directly marginal and reaches only through the adoral half of the initial chamber, which is 0.7 mm long. Septal crowing occurs in specimen FMNH-P30176 between the second to fourth septum with a distance between 0.4 and 0.5 mm, and the subsequent four chambers have a length of 0.7–0.9 mm ( Fig. 29B View Fig ).
Remarks
The assignment of the specimens described above to B. boliviensis is justified herein by the combination of the distinctively pronounced transverse ornamentation, a relatively wide siphuncle, and a relatively large adult size. The relative siphuncular diameter of the Olenidsletta Member specimens is on average slightly wider than that of the holotype and syntype of B. boliviensis . However, the range of measurements of the two types is well within the range of our specimens (see Fig. 24 View Fig ).
Another species of Bactroceras with a similarly wide siphuncle (rSD = ¹/6) is B. wilsoni Flower, 1968 from the Antelope Valley Limestone, Nevada, USA, which differs, however, in lacking the distinctively striated ornamentation. The types of B. angustisiphonatum ( Rüdiger, 1889) differ from the Bactroceras specimens of the Olenidsletta Member in having a very narrow siphuncle with an rSD of ca ¹/12 ( Holm 1898: 9) to ¹/11 ( Rüdiger 1889: 37). Additionally, the ornamentation of B. angustisiphonatum , as originally described by Rüdiger (1889: 37), is weak; Rüdiger (1889) referred to it as “Anwachsstreifen”, i.e., growth lines (this is in contrast to his use of the term “Ringlinien”, i.e., transverse striae, e.g., Rüdiger 1889: 38). The relatively weak ornamentation, with growth lines only, of the original material of B. angustisiphonatum is also explicitly mentioned in Holm (1898: 9). Bactroceras sandbergeri (Barrande, 1867) differs also in having a siphuncle with a diameter of <0.1 of the corresponding conch cross section ( Aubrechtová 2015).
Evans (2005) described an assemblage of ca 40 specimens from the Olenidssletta Member, collected by R.A. Fortey and D.L. Bruton, under “ B. angustisiphonatum (Rüdiger, 1891) ” (sic!). The assemblage came from “the top of division V 2 ” ( Evans 2005: 28), with high probability from exactly the same horizon as most of the specimens assigned to B. boliviensis herein. Bactroceras boliviensis is distinctive within the orthocones of the Olenidssletta Member and occurs in great numbers at 128 m above the base of the member at Profilstranda. The specimens from the Olenidssletta Member described by Evans (2005) are therefore with high probability in large part conspecific with the specimens described herein under B. boliviensis . Our measurements add to the knowledge of the variability of this assemblage, and specifically the dimensions and characters of the apical parts are almost identical (compare Evans 2005: text-fig. 8e). However, B. boliviensis co-occurs with Ethanoceras solitudines gen. et sp. nov., which differs in having a less pronounced ornamentation and a non-marginal, eccentrically positioned siphuncle throughout its entire growth. This makes the distinction between Bactroceras and Ethanoceras difficult or impossible in specimens with diameters <6 mm and without well preserved outer shell or extreme apical parts. Therefore, we measured only specimens with a well-preserved shell surface. Our results differ slightly from the measurements of Evans (2005: text-fig. 8) in having a strictly marginal siphuncle position in B. boliviensis in specimens with diameters>ca 6–7 mm. This difference is potentially a result of unrecognized specimens of Ethanoceras solitudines gen. et sp. nov. in the sample of Evans (2005).
Stratigraphic and geographic range
The types and previously only known specimens of B. boliviensis are from the Pircancha Formation, Bolivia, Baltograptus minutus graptolite zone, which is time equivalent to the lower part of the Didymograptus bifidus graptolite zone (see Gutiérrez-Marco & Martin 2016). The D. bifidus graptolite zone reaches into the upper part of the V 1 division of the Olenidsletta Member ( Cooper & Fortey 1982). The specimens described herein are from V 1 –V 2 trilobite zones; hence the species is known from the Blackhillsian, late Floian of Bolivia and Spitsbergen.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bactroceras boliviensis Aubrechtová, 2015
| Kröger, Björn & Pohle, Alexander 2021 |
Bactroceras boliviensis Aubrechtová, 2015: 206–207
| Aubrechtova M. 2015: 207 |
Bactroceras angustisiphonatum (Rüdiger)
| Evans D. H. 2005: 27 |
Bactroceras aff. avus
| Marek J. & Weber B. & Schonian F. & Egenhoff S. O. & Erdtmann D. 2000: 56 |