
Mantidactylus curtus ( Boulenger, 1882 )
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504338 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FFE9-FFF7-4F13-4D8F4ECB7D6E |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus curtus ( Boulenger, 1882 ) |
| status |
|
Mantidactylus curtus ( Boulenger, 1882) View in CoL
Type material.— Rana curta Boulenger, 1882 is based on seven syntypes, four of which are still available: BMNH 1947.2.10.28–31 from ‘ East Betsileo’ and ‘ Ankafana, Betsileo’ . We here designate the DNA-barcoded specimen BMNH 1947.2.10.30, probably an adult male (sex not unambiguously confirmed), as lectotype. In consequence, the type locality is now ‘ East Betsileo’ . Lectotype designation is justified by the need to stabilize this and other nomina in Brygoomantis , given the uncertain identity and morphological similarity of many taxa in the subgenus.
)
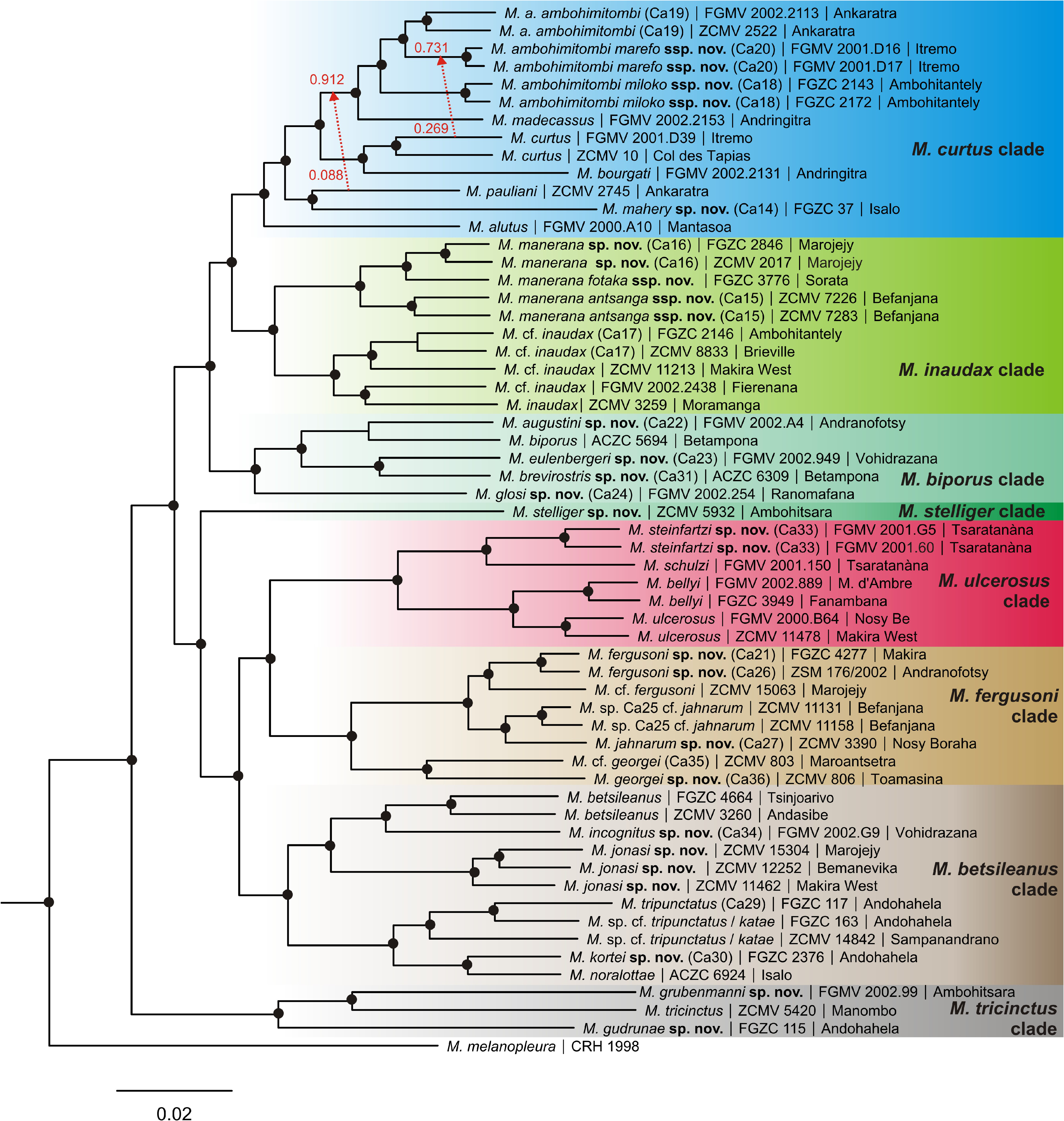
Identity.—The name Mantidactylus curtus has been applied to a complex of genetically divergent lineages inhabiting various mountain ranges and areas of the central high plateau of Madagascar (e.g. Blommers-Schl̂sser & Blanc 1991; Glaw & Vences 1992a). Glaw and Vences (2006) revalidated M. bourgati to refer to the lineage of the Andringitra Massif, but the identity of M. curtus remained uncertain. We here provide a 16S sequence of the lectotype that clusters with a lineage from various localities not far from the type locality (e.g. Antoetra, Itremo, Col des Tapias), providing definitive evidence of the assignment of the nomen curtus to this lineage.
Evidence of introgression of genomic material from this species ( M. curtus ) into a syntopic lineage ( M. ambohimitombi marefo ssp. nov., described below) was found in the Phylonetworks analysis ( Fig. 5 View FIGURE 5 ). Since the latter taxon appears to have a limited distribution range in the Itremo Massif, and the observed reticulation only concerned one M. curtus specimen from the same site, it is likely that this inter-species gene flow is localized and does not compromise the identity of M. curtus as independently evolving lineage.
Synonyms.—Boulenger (1895) considered Rana inaudax Peracca, 1893 to be a synonym of M. curtus , but that species name is revalidated below.
Diagnosis.— A member of the M. curtus clade and sister to M. bourgati . See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of relatively large body size of up to 39 mm, smooth skin, absence of dorsolateral ridges, strongly developed foot webbing with fully webbed fifth toe, and relatively short snout distinguishes this species from species of the other clades.
Within the M. curtus clade, M. alutus , M. madecassus and M. pauliani have smaller body sizes ( Table 4 View TABLE 4 ).
Mantidactylus curtus has smooth dorsal skin, constituting a difference to many specimens of M. ambohimitombi and M. bourgati where the skin is somewhat granular.As far as known, M. curtus and its sister species M. bourgati occur allopatrically and therefore can be distinguished based on localities. For detailed distinction from new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. curtus in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Variation.—Variation in measurements is given in Table 5 View TABLE 5 . See Fig. 10 View FIGURE 10 for colouration in life and its variation.
Evidence for sexual size dimorphism is inconclusive (confirmed male SVL 33.7 mm [n = 1] vs confirmed female SVL 32.5–39.8 mm [n = 8]). In the male specimen MRSN A6757, FGL x FGW is 5.6 mm x 2.6 mm. In many other individuals, femoral glands are less distinct, and some of them cannot be reliably sexed by external examination.
Natural history.— Specimens were found in the vicinity of highland streams, usually quite close to the water.
Calls.—The call of this species has not been recorded.
Tadpoles. —A tadpole of M. curtus (ZSM 943/2004) was described by Schmidt et al. (2009). The tadpole description by Blommers-Schl̂sser (1979), based on materialfromManjakatompo(Ankaratra) andAngavokely, probably refers to different species, as M. curtus is not among the species we have recorded from Ankaratra here. We have not re-sampled Angavokely, but M. alutus and M. ambohimitombi miloko ssp. nov. occur at sites nearby, the latter of which closely resembles M. curtus .
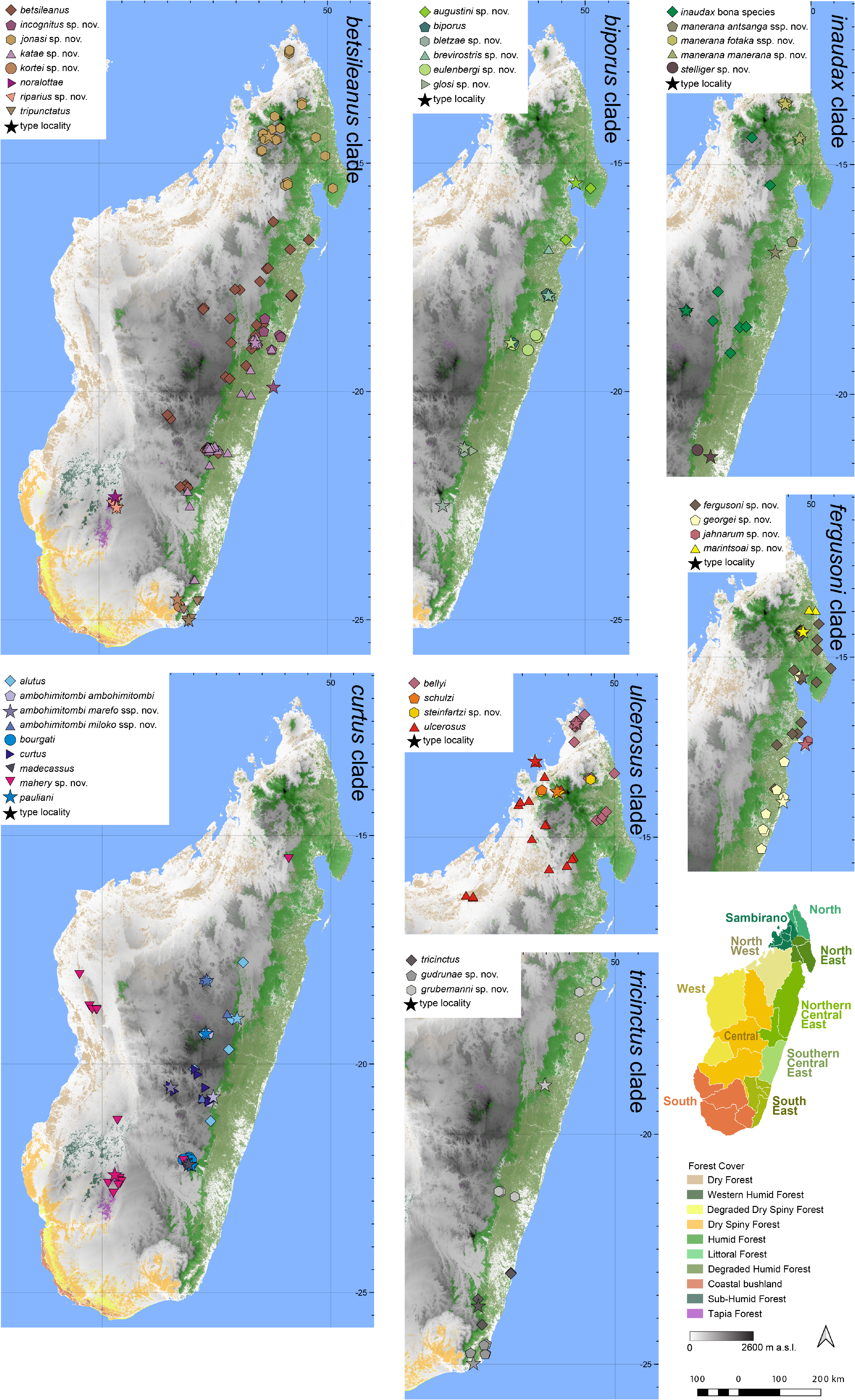
Distribution.— Endemic to a small area of the central highlands of Madagascar ( Fig. 7 View FIGURE 7 ). This species is known from Ambositra, Ankazomivady, Antoetra, Antsirakambiaty forest, Col des Tapias, Ibity, Itremo, and Vatolampy. Elevation range: 1300–2090 m a.s.l.
Etymology.— Latin adjective meaning ‘shortened’ or ‘short’, presumably in reference to the short snout of the species.

| BMNH |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |