
Mantidactylus betsileanus ( Boulenger, 1882 )
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504374 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FFB8-FFA6-4CB1-4C4F49F07C2E |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus betsileanus ( Boulenger, 1882 ) |
| status |
|
Mantidactylus betsileanus ( Boulenger, 1882) View in CoL
Type material.— Rana betsileana Boulenger, 1882 is based on an uncertain number of syntypes ( Boulenger 1882 mentioned seven containers, lettered a–g, but several of these contained more than one specimen), which include BMNH 1947.2.26.33–45 (according to BlommersSchl̂sser & Blanc 1991) and MCZ 15362 (on exchange from BMNH; Barbour & Loveridge 1946), from ‘ East Betsileo’ and ‘ Ankafana , Betsileo’. We here designate BMNH 1947.2.26.45 , an adult male, as lectotype of this species. Lectotype designation is justified by the need to stabilize this and other nomina in Brygoomantis , given the uncertain identity and morphological similarity of many taxa in the subgenus.
Identity.—This nomen has been assigned by Blommers-Schl̂sser (1979) to specimens she collected in the Northern Central East with relatively small femoral glands in males, a typical white tip on the snout, and a single-note, long, pulsed call. This definition of Mantidactylus betsileanus was followed in subsequent accounts (e.g. Glaw & Vences 1992a, 1994, 2007; Vences et al. 2006). Morphology of the lectotype (designated here) and of several paralectotypes examined agrees with this definition. Here we furthermore sequenced by barcode fishing the lectotype BMNH 1947.2.26.45 and paralectotype BMNH 1947.2.26.44, and thus provide genetic confirmation for this assignment.
Synonyms.—Several junior synonyms have been assigned to M. betsileanus (Blommers-Schl̂sser & Blanc 1991; Frost 2021), and the identity of these nomina has remained enigmatic due to the small amount of information on their name-bearing types which are often in a poor state of preservation. By barcode fishing we here robustly assign the following names as junior synonyms to this species:
Rhacophorus fumigatus Mocquard, 1895 , according to Guibé (1950), Blommers-Schl̂sser and Blanc (1991), and Frost (2021) based on holotype MNHN 1895.258 (by monotypy), from ‘ Madagascar.... côte ouest’. The sequence obtained from the holotype specimen clusters firmly among sequences of M. betsileanus View in CoL . It should be noted that the type locality is probably in error as M. betsileanus View in CoL has so far not been found on Madagascar’s western coast.
Mantidactylus multiplicatus Boettger, 1913 View in CoL is based on the holotype (by monotypy) SMF 6733 (formerly 1068.5a) from ‘Alaotra-See, Ost-Madagascar’. The 16S sequence obtained from the holotype firmly clusters with sequences of M. betsileanus View in CoL , confirming this nomen as a junior synonym of M. betsileanus View in CoL , in agreement with previous assertions (e.g. Blommers-Schl̂sser & Blanc 1991). Mantidactylus multiplicatus View in CoL has recently been used as a valid species name (e.g. Poth et al. 2012, 2013) and this view was followed by Frost (2021); however, our new data provide clear evidence that the nomen multiplicatus View in CoL does not apply to the lineage initially studied by Poth et al. (2012), which instead represents a new species, described below as M. katae sp. nov.
Mantidactylus brunneus Ahl, 1929 View in CoL , based on an unnumbered holotype in the ZMB collection from ‘Nord-West-Madagascar’ that has been reported lost ( Frost 2021; Guibé 1978). We rediscovered the holotype of this nomen in the ZMB collection, corresponding to ZMB 30514. The 16S sequence obtained from this specimen firmly clusters with sequences of M. betsileanus View in CoL . As with other species named by E. Ahl, the locality is probably wrong as M. betsileanus View in CoL is not known to occur in northwestern Madagascar.
Another nomen previously considered a synonym of M. betsileanus View in CoL ( Mantidactylus tripunctatus Angel, 1930 View in CoL ) ( Frost 2021) is herein resurrected as the name for a genetically divergent lineage of Brygoomantis from southern Madagascar (see species account below).
Diagnosis.—A member of the M. betsileanus clade as revealed by the phylogenomic analysis, and sister to the poorly known M. incognitus sp. nov. described below. See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of a relatively small body size in males (SVL 22–29 mm) and distinctly larger size in females (SVL 30–37 mm), slightly tubercular dorsal skin with distinct continuous dorsolateral ridges, reduced webbing (one phalanx of fifth toe free of web), absence of white spots on flanks, presence of a white marking on snout tip, and advertisement call consisting of a single, long note composed of more than 100 pulses distinguishes M. betsileanus from species of all other clades. Within the M. betsileanus clade, the species differs from all species except possibly M. incognitus sp. nov. (for which calls are unknown) by a higher number of pulses in its advertisement calls ( Table 4 View TABLE 4 ); it also differs from M. noralottae by smaller size of males, and from the sympatric M. katae sp. nov. by smaller size of femoral glands (see account of that species below). For a detailed distinction from its sister species M. incognitus sp. nov., and from all other new species described herein, see the respective species accounts.A full list of molecular diagnostic sites in the 16S gene of M. betsileanus in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Variation.—Variation in measurements is given in Table 7. See Fig. 34 View FIGURE 34 for colouration in life and its variation. There is moderate sexual size dimorphism (confirmed male SVL 21.9–29.0 mm [n = 11] vs confirmed female SVL 29.5–36.2 mm [n = 10]). Males also have distinctly larger tympanum sizes than females (HTD/ED ratio is 57–78% in females, 72–115% in males). Femoral glands are relatively small, indistinct, and relatively widely spaced, i.e. the distal ulcerous macrogland on opposite thighs are rather widely separated and the proximal granular gland fields are indistinct, although probably present.
The proximal granular gland fields on opposite thighs probably are contacting each other medially (compare with the syntopic M. katae sp. nov. which has much larger femoral glands).
Natural history.—A common species whose typical calls are often heard from small running water bodies in rainforest but also in forest fragments and degraded areas or plantations nearby. Calls are emitted during day and night, especially during the day often from concealed positions directly at the edge of water. Very common in swamp areas, also ricefields next to rainforest, as long as the water is shallow and is at least very slightly flowing.
Usually sitting in shallow water or along the stream bank, hiding in the leaf litter nearby water bodies or sometimes found sitting on low vegetation about 0.3 m hight. In Ranomafana and surrounds found at an elevational range between 550–1132 m a.s.l. M. betsileanus was successfully bred in captivity in the Mitsinjo amphibian husbandry research and captive breeding facility at Andasibe ( Edmonds et al. 2012). The size and structure of a population from near Andasibe was studied by Edmonds et al. (2019).
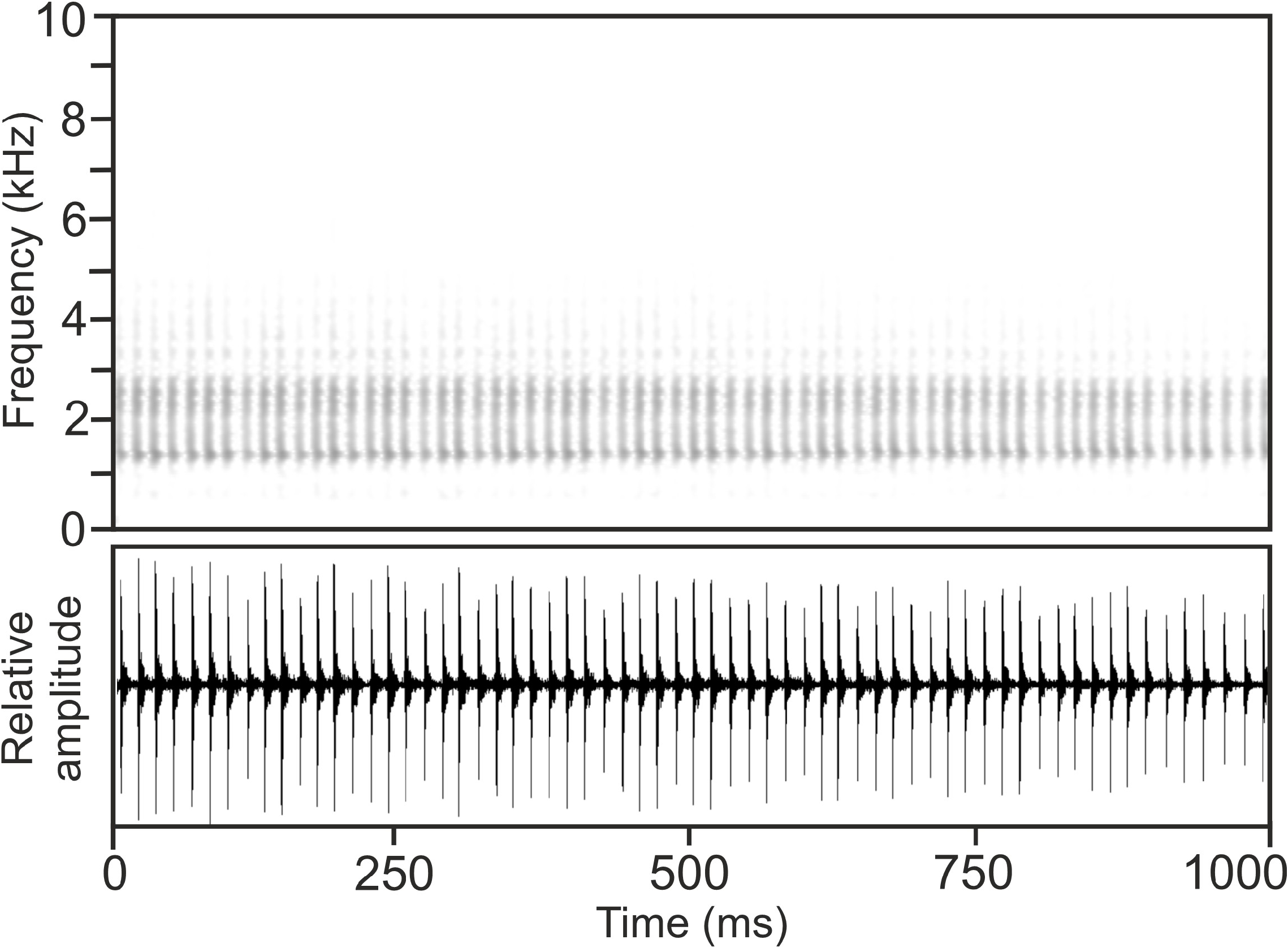
Calls.—The advertisement call of M. betsileanus , recorded on 29 January 1994 at Mandraka, 23–24°C air temperature ( Vences et al. 2006: CD2, track 62, cut 1), consisted of a long, regularly pulsed note ( Fig. 35 View FIGURE 35 ), emitted at more or less regular intervals, but always in slow succession. Notes exhibited some amplitude modulation, with call energy increasing rapidly to the maximum at the beginning of the note, followed by continuous decrease of energy towards the note’s end. Pulse repetition rate was higher in the centre of the note and slightly lower at its beginning and end. Call energy was distributed across a wide frequency range. Numerical parameters of four analysed calls were as follows: call duration (= note duration) 2907–3103 ms (3046.5 ± 93.3 ms); 165–178 pulses per note (173.0 ± 7.7); pulse duration 4–7 ms (5.5 ± 1.0 ms); pulse repetition rate within notes 46.3–64.5 pulses/ s (57.0 ± 5.1); dominant frequency 1399–1421 Hz (1410 ± 12 Hz), with a second peak of almost identical energy at around 2650 Hz; prevalent bandwidth 1100–6000 Hz; call repetition rate (= note repetition rate) ca 3.7 calls/min.
Calls recorded on 12 February 2008 at a forest fragment northeast of Lake Alaotra, at an estimated 20–25°C air temperature, perfectly agreed with the call described above from Mandraka. Numerical parameters of five analysed calls were as follows: call duration (= note duration) 2464–3650 ms (3235.0 ± 438.3 ms); 181– 199 pulses per note (192.3 ± 9.9); pulse duration 4–7 ms (5.3 ± 0.9 ms); pulse repetition rate within notes 39.5–71.4 pulses/s (56.8 ± 12.1); dominant frequency 1356–1464 Hz (1404 ± 42 Hz), with a second peak of almost identical energy at around 2800–2900 Hz; prevalent bandwidth 1100–6000 Hz; call repetition rate (= note repetition rate) ca 3.1 calls/min.
Calls recorded on 25 March 2006, 18:00 h, at the crossing of the Moramanga-Anosibe An’Ala and Besariaka roads, estimated 20–25°C air temperature, also agreed with the calls described from Mandraka. Although difficult to analyse in detail due to many overlapping calls (including those of syntopic M. inaudax ), following numerical parameters could be measured (10 calls originating from at least three individuals analysed): call duration (= note duration) 1560–2279 ms (2010.9 ± 278.6 ms); 107–173 pulses per note (136.0 ± 33.7); pulse duration 2–5 ms (3.6 ± 0.8 ms); pulse repetition rate within notes 63.5–81.6 pulses/s (74.0 ± 6.5); dominant frequency 1261–1574 Hz (1414 ± 148 Hz), with a second peak of almost identical energy at around 2700–3150 Hz; prevalent bandwidth 1000–4200 Hz
Calls recorded on 25 March 2006, at night, at the orchid garden in Andasibe, estimated 20–25°C air temperature, agreed with the calls described above as well. Numerical parameters of six analysed calls, originating from two individuals are as follows: call duration (= note duration) 1887–2868 ms (2439.0 ± 351.9 ms); 173–201 pulses per note (184.3 ± 14.7); pulse duration 4–7 ms (5.5 ± 0.9 ms); pulse repetition rate within notes 56.0–105.3 pulses/s (70.1 ± 17.1); dominant frequency 1238–1340 Hz (1289 ± 38 Hz), with a second peak of almost identical energy at around 2700–2900 Hz; prevalent bandwidth 1000–4600 Hz; call repetition rate (= note repetition rate) ca 4–5 calls/min.
A divergent call has been recorded from specimen MRSN A6343 (FAZC 13875; accession number of 16S sequence HM364713 View Materials ), at Betampona. The calls of this individual were recorded at 19:00 on 14 November 2007, at 20°C air temperature. They consisted of a very long, regularly pulsed note, with very short pulses. Numerical parameters of five analysed calls were as follows: call duration (= note duration) 3490–7507 ms (6148.8 ± 1552.6 ms); 78–164 pulses per note (134.8 ± 33.6); pulse duration 2 ms (2.0 ± 0.0 ms); pulse repetition rate within notes 19.2–24.6 pulses/s (21.9 ± 1.7); dominant frequency 1670–1826 Hz (1719 ± 61 Hz); prevalent bandwidth 1000–4800 Hz. Especially the pulse repetition rate in these calls differed strongly from all other available recordings and we have therefore not included it in the characterization of this diagnostic feature in Table 4 View TABLE 4 .
More data on the advertisement calls and genetics of M. betsileanus at Betampona are necessary to understand the identity of this population and exclude the possibility of mitochondrial introgression into another Brygoomantis species at this site.
Tadpoles.— The tadpole of M. betsileanus was described by Blommers-Schl̂sser (1979), Knoll et al. (2007), and Scheld et al. (2013). The effect of diet on their development was reported by Soamiarimampionona et al. (2015).
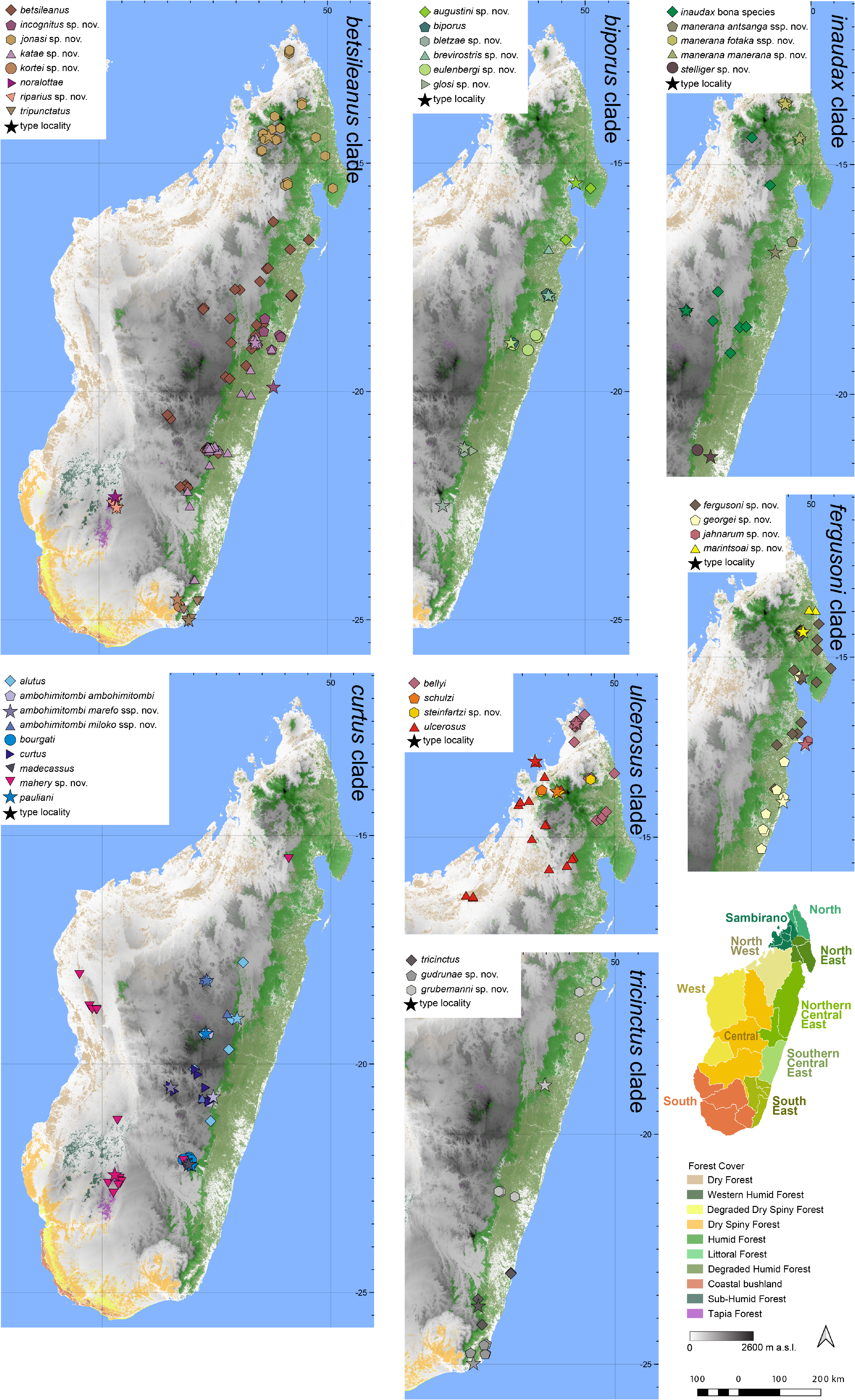
Distribution.— Widespread over a large area of central Madagascar, from sea level on the east coast to the central highlands ( Fig. 7 View FIGURE 7 ). This species is known from the vicinity of Lac Alaotra (type locality of M. multiplicatus ), Ambatovaky, Ambodisakoa, Ambohitantely, Andasibe, Anjozorobe, Anosibe An’Ala, Antara, Antsirakambiaty forest, Befanjana, Betampona, Fierenana, Fivahona, Itremo, Mahasoa, Mandraka, Maromizaha, Marotandrano-Riamalandy, Namoly, Ranomafana and surrounds, Sahambaky Forest (Lakato), Torotorofotsy, Tsaranoro, and Tsinjoarivo. Elevation range: 190–1648 m a.s.l.
Etymology.—Latin adjective meaning ‘of or from Betsileo’, derived from the region Betsileo and the suffix -ânus meaning ‘of or pertaining to’.

| BMNH |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
| MCZ |
USA, Massachusetts, Cambridge, Harvard University, Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mantidactylus betsileanus ( Boulenger, 1882 )
| Scherz, Mark D., Crottini, Angelica, Hutter, Carl R., Hildenbrand, Andrea, Andreone, Franco, Fulgence, Thio Rosin, Köhler, Gunther, Ndriantsoa, Serge Herilala, Ohler, Annemarie, Preick, Michaela, Rakotoarison, Andolalao, Rancilhac, Loïs, Raselimanana, Achille P., Riemann, Jana C., Rödel, Mark-Oliver, Rosa, Gonçalo M., Streicher, Jeffrey W., Vieites, David R., Köhler, Jörn, Hofreiter, Michael, Glaw, Frank & Vences, Miguel 2022 |
M. katae
| Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences 2022 |
Mantidactylus tripunctatus
| Angel 1930 |
Mantidactylus brunneus
| Ahl 1929 |
Mantidactylus multiplicatus
| Boettger 1913 |
Mantidactylus multiplicatus
| Boettger 1913 |
multiplicatus
| Boettger 1913 |
Rhacophorus fumigatus
| Mocquard 1895 |