
Mantidactylus schulzi Vences, Hildenbrand, Warmuth, Andreone & Glaw, 2018
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504365 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF81-FFAF-4F13-4C6F4FF079AE |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus schulzi Vences, Hildenbrand, Warmuth, Andreone & Glaw, 2018 |
| status |
|
Mantidactylus schulzi Vences, Hildenbrand, Warmuth, Andreone & Glaw, 2018 View in CoL
Type material.— Based on holotype (by original designation) ZSM 653/2001 from ‘ Tsaratanàna Massif, Manarikoba Forest , Andampy , “Camp 0” (14.0422°S, 048.7617°E, ca 730 m above sea level), former Antsiranana province, northern Madagascar’. GoogleMaps A total of 19 paratypes: ZMA 19374 ( FGMV 2002.754 ), ZMA 19375 , ZSM 821–826/2003 from Manongarivo, Camp 0 (13.9756°S, 048.4267°E, 688 m a.s.l.); GoogleMaps ZSM 651– 652/2001 and 654/2001 from the type locality; GoogleMaps and the following uncatalogued paratypes from the UADBA collection: FGMV 2002.749 , FGMV 2002.756 , FGMV 2002.758 , FGMV 2002.760 , FGMV 2002.761 , FGMV 2002.763 , FGMV 2002.764 , and FGMV 2002.765 GoogleMaps .
Identity.—This species has been previously referred to as M. sp. 33 ‘Tsaratanàna’ (in Vences et al. (2018) mistakenly stated to be Ca32). It was depicted as ‘ Mantidactylus sp. aff. biporus “Tsaratanàna Andampy”’ by Glaw and Vences (2007). The identity of this smallsized species is well established by genetic data from the holotype and several paratypes provided in the original description.It was previously thought (e.g. Glaw & Vences 2007) to be related to M. biporus , but our phylogenomic tree firmly places it in the M. ulcerosus clade.
Diagnosis.—A member of the M. ulcerosus clade as revealed by the phylogenomic analysis, and sister to the sympatric M. steinfartzi sp. nov. described below. See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of a small body size of up to 29 mm, slightly tubercular dorsal skin, absence of clearly defined, continuous dorsolateral ridges, large tympanum size in males (14–15% of SVL), absence of white spots on flanks, and presence of a white marking on snout tip, distinguishes M. schulzi from species of most other clades exept for the M. betsileanus clade and M. fergusoni clade. It differs from members of the M. fergusoni clade by smaller body size of both sexes, and from members of the M.betsileanus clade by smaller body size of females, and by a combination of more expressed webbing and larger femoral glands ( Table 4 View TABLE 4 ), and from all other Brygoomantis with known advertisement calls by details of temporal call variables. Within the M. ulcerosus clade, M. schulzi differs by a distinctly smaller body size and several other characters from M. bellyi and M. ulcerosus . For a detailed comparison with its sister species M. steinfartzi sp. nov., see description of that species below; for detailed distinction from other new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. schulzi in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Variation.—Variation in measurements is given in Table 6 View TABLE 6 . See Fig.29 View FIGURE 29 for colouration in life and its variation.
There is moderate sexual size dimorphism (confirmed male SVL 21.4–23.5 mm [n = 3] vs confirmed female SVL 24.7–28.9 mm [n = 7]). Males have a somewhat larger tympanum than females (HTD/ED ratio is 71– 82% in females, 91–94% in males). Femoral glands are very prominent in males, with a particularly large distal ulcerous macrogland, and a relatively small proximal granular gland field; the granular gland fields on the two opposite thights contact each other medially. The glands have a light brown to yellowish tone in life, but are not distinctly orange as in the large-sized species M. bellyi and M. ulcerosus .
Natural history.—All specimens were observed in small streams and rivulets in primary rainforest. Calling males were observed directly next to the water. Both in Tsaratanàna and Manongarivo, this species occurred at slightly lower elevation than its sister species, M. steinfartzi sp. nov., but the sites are in close proximity to one another (i.e. <500 m linear distance at Manongarivo, <2600 m at Tsaratanàna).
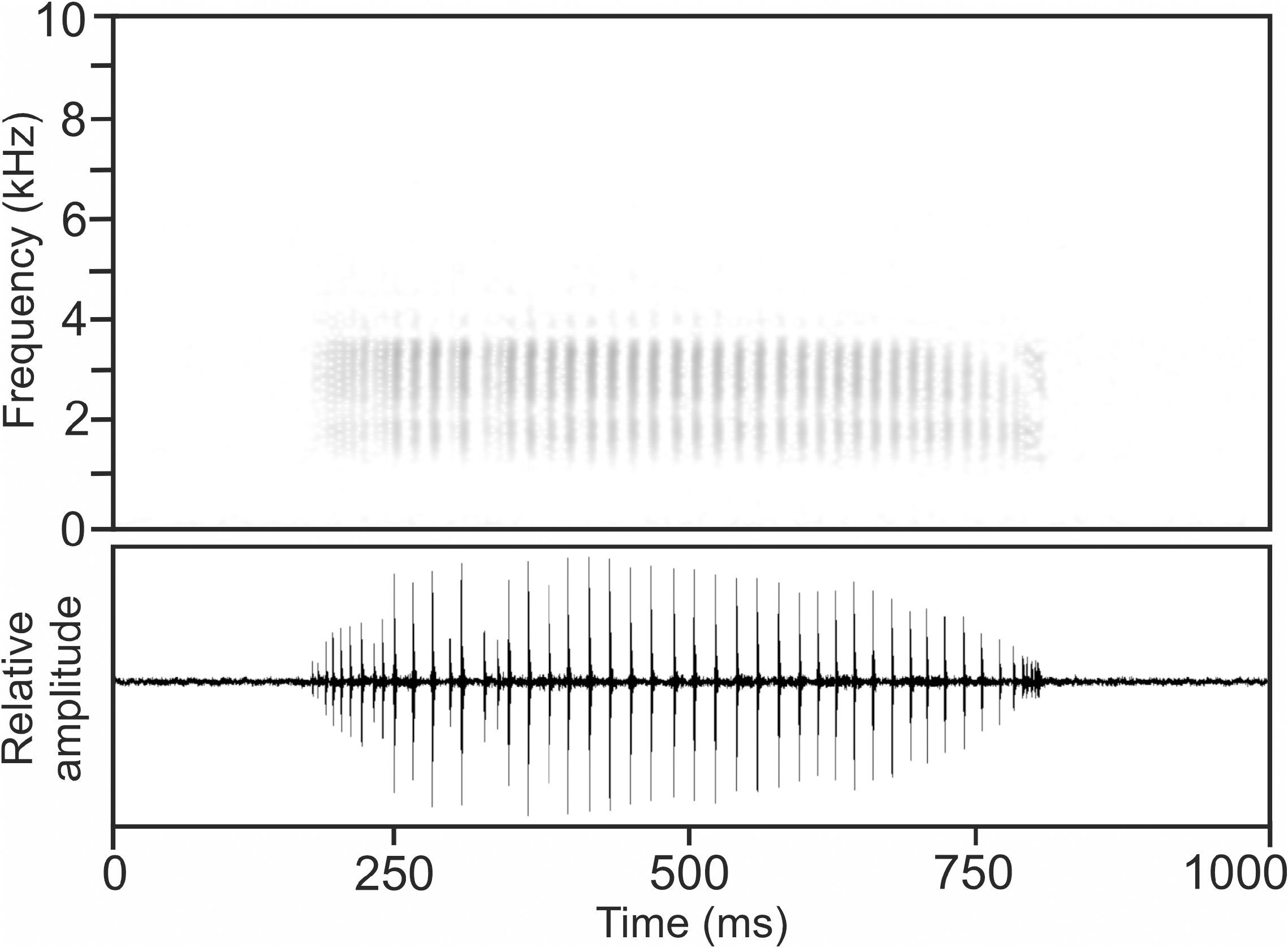
Calls.— The advertisement call of M. schulzi recorded on 10–11 February 2001 at Andampy campsite, Manarikoba forest, Tsaratanàna Strict Nature reserve, 25– 26°C air temperature ( Vences et al. 2006: CD2, track 70), has been adequately described by Vences et al. (2018). It consists of a pulsed note of very variable duration ( Fig. 30 View FIGURE 30 ), sometimes emitted in short series of 2–3 calls. Numerical parameters of 30 analysed calls are as follows: call duration (= note duration) 11–996 ms (301 ± 306 ms); 6–73 pulses per note (27 ± 21); pulse duration 1–3 ms (2 ± 1 ms); pulse repetition rate within notes approximately 55–130 pulses/s; dominant frequency is difficult to determine, but roughly ranges between 2900–3400 Hz; prevalent bandwidth 1500–3900 Hz; call repetition rate (= note repetition rate) within regular series ca 30–50 calls/min.
Tadpoles.— The tadpole of this species has not been described.
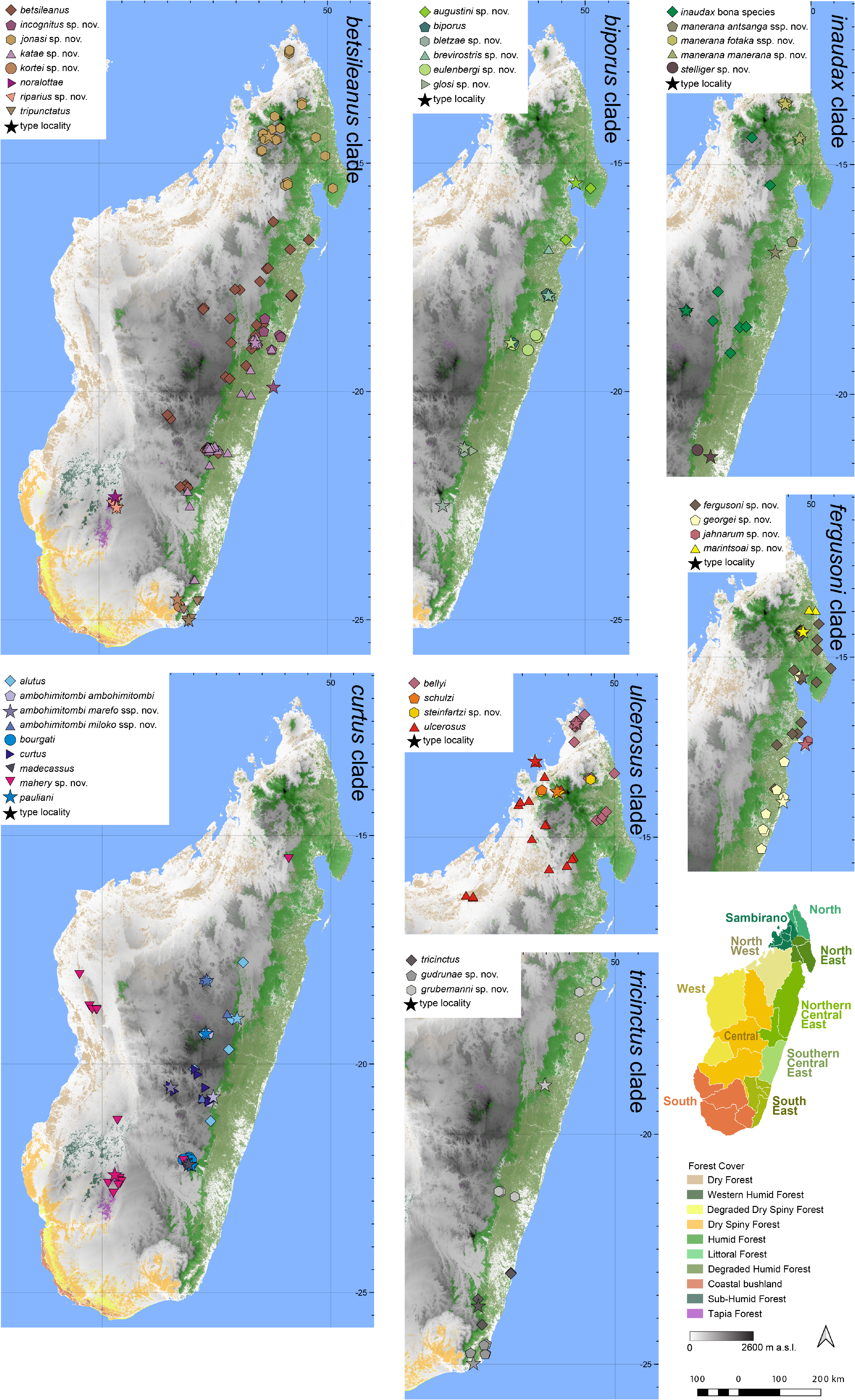
Distribution.— Endemic to the Sambirano Region in northern Madagascar ( Fig. 7 View FIGURE 7 ). This species is known from Tsaratanàna (Manarikoba, type locality)and Manongarivo. Elevation range: 688–730 m a.s.l.
Etymology.— Eponym for Stefan Schulz, professor of organic chemistry at Braunschweig University of Technology, in recognition of his contributions to the study of volatile pheromones in Brygoomantis and other mantellines.

| ZSM |
Germany, Muenchen [= Munich], Zoologische Staatssammlung |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |